Biologie pro Obory
Výsledky Učení
- Popsat úlohu RNA polymeráza
Tato část se bude rozšiřovat na konkrétní úlohu RNA polymerázy během transkripce. Čtěte dále a dozvíte se roli RNA polymeráz v každé fázi transkripce.,
Zahájení Transkripce
na Rozdíl od prokaryotické polymeráza, který se může vázat na DNA šablony, na jeho vlastní, eukaryota vyžadují několik dalších proteinů, tzv. transkripční faktory, první se váží na promotorové oblasti a pak pomoci získávat příslušné polymerázy.
Tři Eukaryotické RNA Polymerázy
funkce eukaryotické mRNA syntézy jsou výrazně složitější těchto prokaryot. Místo jediné polymerázy obsahující pět podjednotek mají eukaryoty tři polymerázy, z nichž každá tvoří 10 podjednotek nebo více., Každá eukaryotická polymeráza také vyžaduje zřetelnou sadu transkripčních faktorů, které ji přivedou do šablony DNA.
RNA polymeráza I se nachází v jadérko, specializované jaderné spodku, ve kterém ribozomální RNA (rRNA) je přepsán, zpracovány a sestaveny do ribozomy (Tabulka 1). Molekuly rRNA jsou považovány za strukturální RNA, protože mají buněčnou roli, ale nejsou přeloženy do bílkovin. RRNA jsou součástí ribozomu a jsou nezbytné pro proces překladu. RNA polymeráza I syntetizuje všechny rRNA s výjimkou molekuly 5S rRNA., Označení “ S „se vztahuje na jednotky“ Svedberg“, nonadditive hodnota, která charakterizuje rychlost, při které částice sedimenty během odstředění.
| Tabulka 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Eukaryotické pre-mRNA procházejí rozsáhlým zpracováním po transkripci, ale před překladem (Obrázek 1). Pro přehlednost bude diskuse tohoto modulu o transkripci a překladu v eukaryotách používat termín „mRNA“ k popisu pouze zralých, zpracovaných molekul, které jsou připraveny k překladu. RNA polymeráza II je zodpovědná za přepis drtivé většiny eukaryotických genů.

Obrázek 1. Eukaryotická mRNA obsahuje introny, které musí být spojeny. Přidá se také 5′ cap a 3′ poly-a ocas.,
RNA polymeráza III je také umístěna v jádru. Tato polymeráza přepisuje řadu strukturních RNA, která zahrnuje 5S pre-rRNA, transfer pre-RNA (pre-tRNA) a Malé jaderné pre-RNA. TRNA mají zásadní roli v překladu; slouží jako adaptorové molekuly mezi šablonou mRNA a rostoucím polypeptidovým řetězcem. Malé jaderné RNA mají řadu funkcí, včetně“ spojování “ pre-mRNA a regulace transkripčních faktorů.,
vědec charakterizující nový gen může určit, které polymeráza přepisuje to tím, že testování, zda je gen exprimován v přítomnosti určitého houby jed, α-amanitin (Tabulka 1). Zajímavé je, že α-amanitin produkován Amanita phalloides, Smrt Čepice houby, ovlivňuje tři polymerázy velmi odlišně. RNA polymeráza I je zcela necitlivá na α-amanitin, což znamená, že polymeráza může přepsat DNA in vitro za přítomnosti tohoto jedu. Naproti tomu RNA polymeráza II je extrémně citlivá na α-amanitin a RNA polymeráza III je mírně citlivá., Znalost přepisování polymerázy může vést výzkumníka k obecné funkci studovaného genu. Protože RNA polymeráza II přepisuje převážnou většinu genů, zaměříme se na tuto polymerázu v našich následných diskusích o eukaryotických transkripčních faktorech a promotorech.
promotory RNA polymerázy II a transkripční faktory
eukaryotické promotory jsou mnohem větší a složitější než prokaryotické promotory. Oba však mají sekvenci podobnou sekvenci prokaryot -10., V eukaryotách se tato sekvence nazývá TATA box a má konsensuální sekvenci TATAAA na kódovacím řetězci. Nachází se na bázi -25 až -35 vzhledem k místu zahájení (+1) (Obrázek 2). Tato sekvence není totožná s krabicí E. coli -10, ale zachovává prvek bohatý na A–T. Termostabilita vazeb A-T je nízká, což pomáhá šabloně DNA lokálně uvolnit při přípravě transkripce.,
Namísto jednoduché σ faktor, který pomáhá vázat prokaryotické RNA polymeráza svého zřizovatele, eukaryota sestavit komplexní transkripční faktory potřebné pro nábor RNA polymeráza II protein kódujících genů. Transkripční faktory, které se váží na promotor, se nazývají bazální transkripční faktory. Tyto bazální faktory se nazývají TFII (pro transkripční faktor/polymerázu II) plus další písmeno (A-J). Jádrem komplexu je tfiid, který zahrnuje protein vázající TATA (TBP)., Další transkripční faktory systematicky spadají do místo na templátu DNA, s každou další stabilizaci preiniciačního komplexu a přispět k náboru RNA polymerázy II.
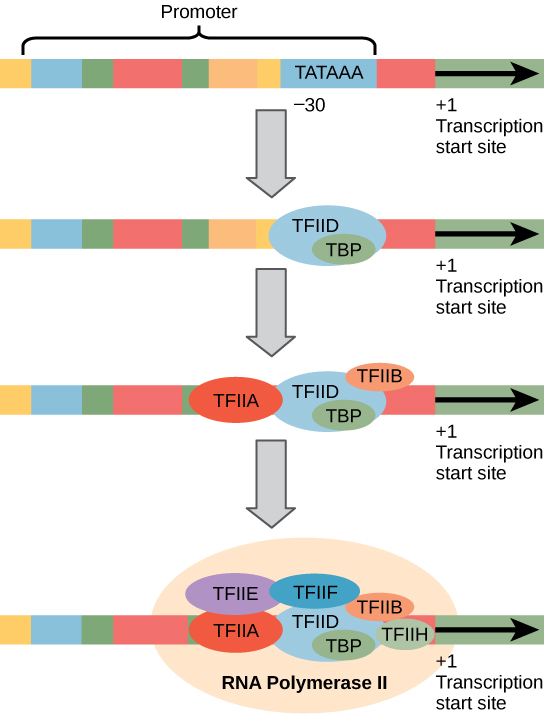
Obrázek 2. Je zobrazen generalizovaný promotor genu transkribovaného RNA polymerázou II. Transkripční faktory rozpoznávají promotor. RNA polymeráza II se pak váže a tvoří komplex iniciace transkripce.,
Cvičení Otázku
vědec spojek a eukaryotický promotor před bakteriální gen a vloží genu bakteriální chromozom. Čekali byste, že bakterie přepisují Gen?
některé eukaryotické promotory mají také konzervovaný CAAT box (GGCCAATCT) přibližně -80. Dále před boxem TATA mohou propagátoři eukaryotic obsahovat také jednu nebo více krabic bohatých na GC (GGCG) nebo oktamerové krabice (ATTTGCAT)., Tyto prvky vázat buněčné faktory, které zvyšují účinnost transkripce iniciace a jsou často identifikovány ve více „aktivní“ geny, které jsou neustále vyjádřené buňky.
bazální transkripční faktory jsou rozhodující pro vytvoření preinitačního komplexu na šabloně DNA, který následně rekrutuje RNA polymerázu II pro zahájení transkripce. Složitost eukaryotické transkripce nekončí polymerázami a promotory., Armáda dalších transkripčních faktorů, které se váží na zesilovače proti proudu a tlumiče hluku, také pomáhají regulovat frekvenci, s níž je pre-mRNA syntetizována z genu. Zesilovače a tlumiče hluku ovlivňují účinnost transkripce, ale nejsou nutné, aby transkripce pokračovala.
vývoj promotorů
vývoj genů může být známým konceptem. Mutace se mohou objevit v genech během replikace DNA a výsledek může nebo nemusí být pro buňku prospěšný., Změnou enzymu, strukturního proteinu nebo jiného faktoru může proces mutace transformovat funkce nebo fyzikální vlastnosti. Mohou se však vyvinout i eukaryotické promotory a další genové regulační sekvence. Zvažte například gen, který se po mnoho generací stává pro buňku cennější. Možná gen kóduje strukturální protein, který buňka potřebuje k syntéze v hojnosti pro určitou funkci. Pokud tomu tak je, bylo by pro buňku prospěšné, aby promotor tohoto genu účinněji přijímal transkripční faktory a zvyšoval genovou expresi.,
vědci zkoumající vývoj promotorových sekvencí hlásili různé výsledky. Částečně je to proto, že je obtížné přesně odvodit, kde začíná a končí eukaryotický promotor. Některé promotory se vyskytují v genech; jiné jsou umístěny velmi daleko proti proudu nebo dokonce po proudu genů, které regulují. Když však vědci omezili své vyšetření na sekvence promotorů lidského jádra, které byly experimentálně definovány jako sekvence, které vážou preinitační komplex, zjistili, že promotory se vyvíjejí ještě rychleji než geny kódující proteiny.,
stále není jasné, jak by promotorová evoluce mohla odpovídat vývoji lidí nebo jiných vyšších organismů. Vývoj promotoru k účinnému vytváření více či méně daného genového produktu je však zajímavou alternativou k vývoji samotných genů.
Pořadatel Struktury RNA Polymerázy i a III
procesy přináší RNA polymerázy i a III k templátu DNA zapojit o něco méně komplexní sbírky transkripčních faktorů, ale obecně téma je stejné.,
zachovaných pořadatel prvky pro geny přepsal polymerázy i a III se liší od těch, přepisována pomocí RNA polymerázy II. RNA polymeráza I přepisuje geny, které mají dva GC-bohaté promotor sekvence v -45 až +20 regionu. Tyto sekvence samotné jsou dostatečné pro zahájení transkripce dochází, ale pořadatelé s další sekvence v oblasti od -180 do -105 před zahájením stránky dále zvýší zahájení. Geny, které jsou transkribovány RNA polymerázou III, mají upstream promotory nebo promotory, které se vyskytují v samotných genech.,
eukaryotická transkripce je přísně regulovaný proces, který vyžaduje interakci různých proteinů mezi sebou a s řetězcem DNA. I když proces transkripce u eukaryot zahrnuje větší metabolické investice než u prokaryot, to zajišťuje, že buňka přepisuje přesně pre-mRNAs, že to potřebuje pro syntézu bílkovin.,
Prodloužení a Ukončení
Po vytvoření preinitiation komplex, polymerázy se uvolní z jiné transkripční faktory, a prodloužení je povoleno pokračovat tak, jako je tomu u prokaryot s polymeráza syntetizující se pre-mRNA v 5′ 3′ směr. Jak již bylo zmíněno dříve, RNA polymeráza II přepisuje hlavní podíl eukaryotických genů, takže tato část se zaměří na to, jak tato polymeráza dosahuje prodloužení a ukončení.,
ačkoli enzymatický proces prodloužení je v podstatě stejný u eukaryot a prokaryot, šablona DNA je složitější. Když se eukaryotické buňky nerozdělují, jejich geny existují jako difúzní hmotnost DNA a proteinů nazývaných chromatin. DNA je pevně zabalena kolem nabitých histonových proteinů v opakovaných intervalech. Tyto komplexy DNA-histonu, souhrnně nazývané nukleosomy, jsou pravidelně rozmístěny a zahrnují 146 nukleotidů DNA navinutých kolem osmi histonů jako nit kolem cívky.,
aby došlo k syntéze polynukleotidů, musí transkripční zařízení přesunout histony z cesty pokaždé, když narazí na nukleosom. Toho je dosaženo speciálním proteinovým komplexem zvaným fakt, který znamená „usnadňuje transkripci chromatinu.“Tento komplex táhne histony od šablony DNA, když se polymeráza pohybuje podél ní. Jakmile je pre-mRNA syntetizována, komplex faktů nahrazuje histony, aby znovu vytvořil nukleosomy.
ukončení transkripce je pro různé polymerázy odlišné., Na rozdíl od prokaryot, prodloužení pomocí RNA polymerázy II v eukaryot probíhá 1,000–2,000 nukleotidů za koncem gen je přepsán. Tento ocas před mRNA je následně odstraněn štěpením během zpracování mRNA. Na druhé straně RNA polymerázy I a III vyžadují terminační signály. Geny transkribované RNA polymerázou i obsahují specifickou 18-nukleotidovou sekvenci, která je rozpoznána terminačním proteinem. Proces ukončení RNA polymerázy III zahrnuje mRNA vlásenku podobnou Rho-nezávislému ukončení transkripce v prokaryotách.,
zkuste to
Přispějte!
Vylepšete tuto stránku více
- H Liang et al.,“ Rychlý vývoj hlavních promotorů v primátových genomech“, molekulární biologie a evoluce 25 (2008): 1239-44. ↵















