Frontiers in Plant Science (Čeština)
Úvod
Správný průběh zrání ovoce je nezbytné pro dosažení optimální kvality ovoce a dlouhou trvanlivost, je důležité rysy, které určují cenu trhy a konečné zisky. Nicméně, zrání je složitý proces, kde působí řada faktorů, včetně hormonální regulace, která reguluje biochemické a fyziologické změny, které dávají finální organoleptické a nutriční vlastnosti ovoce., Tradičně, masité plody byly klasifikovány podle tam zrání procesu do klimakterických a non-klimakterických plodů. Klimakterické plody, jako jsou rajčata nebo banány vyžadovat nárůstu rychlost dýchání a výroba ethylenu, aby se uvolnil procesu zrání v autokatalytická reakce (Barry a Giovannoni, 2007; Liu et al., 2015)., Antagonistically, non-klimakterický ovoce, jako jsou citrusové plody nebo hrozny nevykazují výbuch ani v ethylenu, ani dýchání před nástupem zrání, a v těchto plodů, zrání je řízena hlavně progresivní akumulace fytohormonů kyseliny abscisové (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Nicméně, nové studie posiluje myšlenku, že proces zrání je nejen řídí produkce fytohormonů, ale spíše se zdálo, že výsledek řízené hormonální rovnováhu (Symons et al., 2012; Teribia et al., 2016; Li et al., 2019).,
auxiny, kyselina salicylová (SA) a melatonin jsou fytohormony zapojené do signalizace a regulace mnoha klíčových procesů v rostlinách. Auxiny byly široce popsány jako regulátory růstu a vývoje s více funkcemi v rostlinách (viz Taylor-Teeples et al., 2016); SA spouští obrannou reakci proti biotrofickým a hemi-biotrofickým patogenům (Loake a Grant, 2007) a hraje důležitou roli při abiotickém stresu (Dong et al., 2014; Wani et al., 2017), řízení kvetení a buněčného cyklu (Carswell et al., 1989; Eberhard et al.,, 1989, resp); a melatonin bylo nejen zjištěno, že mají auxin podobné funkce (Chen et al., 2009; Zuo et al., 2014; Wen et al., 2016), ale to také bylo navrhl, aby působit jako potenciální antioxidant v některých rostlinách (Arnao a Hernández-Ruiz, 2015) a regulátor rostlina odpovědi na patogeny (Chen et al., 2018). Je zajímavé, že tyto tři hormony mají společný prekurzor – chorismate -, tedy metabolické cross-talk dochází mezi nimi, a počet genů musí být jemně regulovaný odklonit chorismate metabolismus směrem těchto sloučenin., Během posledních několika let se několik recenzí zabývalo biosyntézou a rolí auxinu (Paul et al., 2012), SA (Asghari a Aghdam, 2010) a melatonin (Feng et al., 2014; Arnao a Hernández-Ruiz, 2018) v ovoci. Jejich biosyntéza a funkce však byly většinou popsány samostatně. Zde budeme diskutovat společné a rozdílné aspekty v biosyntéze, metabolismu a funkci auxinu, SA a melatoninu na růst a zrání klimakterických a non-klimakterický ovoce., Důraz bude kladen na metabolické odklon klíčové body v jejich biosyntéza z chorismate, regulační úlohu během zrání klimakterických a non-ovoce menopauza, a jejich role v modulaci biosyntézy biologicky aktivních látek, které do značné míry určuje kvalitu ovoce.
fytohormony odvozené od Chorismátu
Chorismát je konečným produktem shikimátové dráhy a hraje klíčovou roli v biosyntéze fytohormonů (Obrázek 1)., Chorismate dává vzniknout aromatických aminokyselin, včetně tryptofanu, a přes několik reakcí, včetně přeměny tryptofanu na indol-3-pyrohroznová kyseliny tryptofan aminotransferázy (TA); to může vytvářet auxin je indol-3-octové (IAA). TA byla navržena jako univerzální klíčový enzym biosyntézy IAA nejen ve vegetativních orgánech (Enríquez-Valencia et al., 2018), ale také ve vývoji reprodukčních orgánů (Reyes-Olalde et al., 2017), včetně růstu a zrání ovoce (Estrada-Johnson et al., 2017)., Naproti tomu, když je aktivována isochorismátsyntáza (ICS), chorismát může být přeměněn na isochorismát a ten může být přeměněn na SA. Navzdory skutečnosti, že tato cesta byla poprvé identifikována v bakteriích, je také v současné době dobře zavedena v rostlinách (Wildermuth et al., 2001; Uppalapati et al., 2007; Catinot et al., 2008; Garcion et al., 2008; Abreu a Munné-Bosch, 2009). Nakonec se tryptofan může změnit na tryptamin enzymem tryptofan dekarboxyláza (TDC)., I když je to další z více tras pro biosyntézu IAA (tryptamin změní na indol-3-acetaldehyd a později do IAA), TDC bylo také navrženo být první, rate-limiting enzymem v melatonin dráhy (Kang et al., 2007). TDC byl poprvé identifikován v rodině Apocynaceae (De Luca et al., 1989), které mají být později popsány v řadě rostlinných systémů (Byeon et al., 2012, 2014; Zhao et al., 2013; Wei et al., 2018)., Po dekarboxylaci, tryptamin je přeměněn na serotonin, které tryptamin-5-hydroxylázy, a pak serotoninu transformuje na N-acetyl-serotoninu že se konečně výnos melatoninu v cytosolu (Obrázek 1).
Obrázek 1. Biosyntéza kyseliny indol-3-octové (IAA), kyseliny salicylové (SA) a melatoninu (Mel) z chorismátu. Všechny tři sloučeniny sdílejí společný konečný prekurzor (chorismát, na druhou oranžově) z shikimátové dráhy., Hlavní body odklonu jsou označeny červenými šipkami a odpovídající enzymy jsou očíslovány v sestupném pořadí divergence od shikimátu. Tučné přerušované šipky označují více než jeden krok mezi sloučeninami. Tenké přerušované šipky označují cesty, které stále nejsou dobře definovány.
klíčové enzymy, které vedou k syntéze hormonů chorismátové dráhy, byly nedávno identifikovány v masitých plodech (Obrázek 1). ICS, který výraz se ukázal být rozhodující pro biotickou stresovou toleranci (Garcion et al.,, 2008; Zeng a on, 2010), byl identifikován v rajčatech a jablkách. Zhu et al. (2016) hlášeny nadměrné exprese ICS 1 (isochorismate syntáza 1) za chladných podmínek skladování v rajčatech, zatímco Zhang et al. (2017) našel aktivaci transkripčního faktoru zapojeného do signální dráhy SA související s patogenem vyvolávající expresi ICS v jablkách. Zvýšená aktivita TDC byla hlášena u nezralých plodů pepře po infekci patogeny-zvýšením exprese TDC1 a TDC2 (Park et al., 2013) – a ve fázi růstu moruše ovoce (Wang et al., 2016)., Nakonec byla TA identifikována v grapevine, a to jak v pre -, tak veraison (Böttcher et al., 2013; Gouthu a Deluc, 2015), stejně jako během zrání jahod (Estrada-Johnson et al., 2017). Je však třeba poznamenat, že geny a enzymy popsané v klimakterických plodech (tj. ICS) nebyly identifikovány v ne-klimakterických plodech (tj.,
Role Chorismate Odvozené od Hormonů v období Menopauzy a Non-Ovoce Menopauza
Odhalují mechanismy ovoce rozvoj byl jednou z hlavních výzev v posledních agronomického výzkumu pro jeho ekonomické důsledky. V této souvislosti byly fytohormony zdůrazněny jako odpovědné hybatele dozrávání ovoce, zejména ethylenu a ABA v klimakterických a ne-klimakterických plodech. To, že tyto fytohormony mohly regulovat samotný vývoj ovoce, se však brzy ukázalo jako příliš jednoduché., Po rozsáhlém výzkumu a se zlepšením analytické chemie a molekulárních technik bylo několik dalších hormonů potvrzeno jako potenciální regulátory vývoje a zrání ovoce, včetně fytohormonů odvozených od chorismátu.
Auxiny Cross-Talk S Jinými Hormony Během Ovocné Sady, Růst a Zrání
Auxiny jsou skupinou rostlinných hormonů, které hrají zásadní roli v vývoj plodu, jak se projevují jejich vlastní vliv a moduluje expresi jiných fytohormonů., Endogenní obsah IAA jsou obzvláště vysoké na ovocné sady a během počáteční růst vývojové fáze, po které IAA množství tendenci klesat před nástupem zrání, a to jak v období menopauzy (Zaharah et al., 2012) a non-klimakterické ovoce (Symons et al., 2012; Teribia et al., 2016), zřejmě až na některé výjimky, jako broskve (Tatsuki et al., 2013) a některé odrůdy švestek (El-Sharkawy et al., 2014; obrázek 2a). Bylo prokázáno, že IAA se podílí na ovoce nastavit zahájení v kombinaci s gibereliny (Mezzetti et al., 2004; Serrani et al., 2010; Bermejo et al., 2018; Hu et al.,, 2018). Poškození biosyntézy nebo signalizace IAA obecně vede k parthenokarpii ovoce, i když to může také vést k abnormálnímu zrání u některých plodů (Wang et al., 2005; Liu J.et al., 2018; Reig et al., 2018). Vysoký obsah IAA v počátečních fázích vývoje ovoce podporovat ovoce růst vzhledem k auxinu důsledky v buněčné dělení v kombinaci s cytokininy a v kontrole buněčné expanze v kombinaci s gibereliny (Liao et al., 2018)., Během tohoto období, hormonální přeslechy mezi auxiny a gibereliny navíc umožňuje normální ovoce tvarování v vyladěn nařízení zprostředkované Auxin Response Factors (ARFs; Liao et al., 2018; Liu s.et al., 2018).
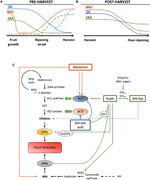
Obrázek 2. Role IAA, SA a Mel během vývoje klimakterických a ne-klimakterických plodů. Model shrnující interakce IAA, SA, a Mel v průběhu zrání klimakterických a non-ovoce menopauza, během (A) před a (B) po sklizni., Přerušované čáry naznačují alternativní dynamiku obsahu fytohormonu v některých plodech (Viz text k diskusi). C) přehled interakcí biosyntézy IAA, SA a Mel s ethylenem a kyselinou abscisovou (ABA) v klimakterických a neklimakterických plodech. Auxin je pozitivním regulátorem biosyntézy ethylenu aktivací genů ACC syntázy (ACS) a indukcí exprese faktorů reagujících na ethylen (ERFs). Auxin také potlačuje produkci ABA represí 9-CIS-epoxykarotenoidní dioxygenázou (NCED) a inhibicí stresových genů reagujících na ABA (ARS) proteinem ARF2., Endogenní obsah IAA může být snížen bílkovinami GH3, jejichž syntéza je podporována ethylenem, ABA a cukry. Diskontinuální čára označuje možnou roli IAA IAA-Asp jako faktoru zrání, který zvyšuje ethylen v některých plodech. Melatonin zvyšuje produkci ABA a ethylenu, ačkoli ACS může být inhibována v závislosti na obsahu melatoninu. SA působí jako inhibitor ethylen potlačující ACC oxidázy (ACO) genů, i když může podporovat expresi ACS., MTA, 50-methylthioadenosin; s-AdoMet, S-adenosyl-methionin; SAM syntáza, S-adenosyl-methionin syntáza; ACC, 1-aminocyklopropan-1-karboxylová kyselina; IPP, isopentenyldifosfát.
snížení obsahu endogenního IAA bylo hlášeno před začátkem zrání v několika plodech. Tato snížení souvisela s konjugací IAA s kyselinou asparagovou (IAA-Asp) syntetázami IAA-amido, GH3-1 a GH3-2 (Obrázek 2c)., GH3-1 a GH3 – 2 skutečně vykazovaly vyšší expresi během raného vývoje ovoce a zejména během zahájení zrání jak u klimakterických plodů, jako jsou rajčata (Sravankumar et al., 2018) a jablka (Onik et al., 2018), stejně jako v ne-klimakterických plodech, jako jsou hroznové bobule (Böttcher et al., 2010, 2011) a maliny (Bernales et al., 2019)., Zajímavé je, že hroznové bobule vykazovaly zvýšenou expresi GH3-1 po aplikaci ABA a ethefonu, což by mohlo vysvětlit zapojení ethylenu do kontroly obsahu IAA po nástupu zrání, a to i v ne-klimakterických plodech (Böttcher et al., 2010). Ve skutečnosti, několik studií zvýraznit těsné interakce mezi auxiny a ethylenu při zrání ovoce, vzájemný vliv mezi nimi (Tadiello et al., 2016a; Busatto et al., 2017)., Pro ovoce menopauza, zvýšený obsah IAA jsou nezbytné pro aktivaci exprese ACC syntázy geny (ACS), kódování ACC syntázu, který, podle pořadí, povede k ethylenu produkce spuštění procesu zrání (Obrázek 2C; Tatsuki et al., 2013). Ovoce jako rajčata (Li et al., 2016) nebo banány (Choudhury et al.,, 2008), které obvykle zkušenosti snížena IAA obsah před nástupem zrání, ukazují zrání zpoždění při IAA nebo analogy jsou exogenně aplikován, zatímco ovoce jako broskve, jejichž obsah IAA postupně zvyšovat, dokud se plně dozrát, show, zrychlené zrání, kdy auxiny jsou aplikovány (Tadiello et al., 2016b). Proto musí existovat těsná, složitá a diferenciální regulace interakcí auxin-ethylen v různých plodech během předhlazení zrání., Kromě toho, IAA obsah pokles před nástupem zrání v některých non-ovoce menopauza, aby de-potlačování 9-cis-epoxycarotenoid dioxygenase (a když už prohlásil) a začít syntézu ABA (Obrázek 2C; Jia et al., 2016). Je zajímavé, že tento proces je také zprostředkován konjugací IAA prostřednictvím zvýšené aktivity GH3 – 1 (Böttcher et al., 2010). Nicméně, Estrada-Johnson et al. (2017), ukázala, že, i když IAA obsah jahodový pokles v průběhu zrání, výraz FaAux/IAA a FaARF genové rodiny jsou vyvolané v červené nádoby, což naznačuje zapojení auxinu signalizace v plně dozrávají plody.,
Během post-sklizni, auxin obsah obvykle zůstávají invariantní nebo mají tendenci klesat v důsledku oxidačních procesů, které mohou vést k malé, ale progresivní snížení endogenní obsah IAA (Obrázek 2B). Auxin ošetření po sklizni zpoždění over-zrání v některých plodů (Chen et al ., 2016; Moro et al., 2017) a zvýšit obsah některých organických kyselin, udržovat kyselost ovoce (Li et al., 2017), což naznačuje, že auxin také hraje významnou roli při kontrole dozrávání ovoce během po sklizni.,
Více Rolí Salicylové Kyseliny V Ovoce Vývoj a Zrání
kyselina Salicylová je další chorismate-odvozené fytohormonů, které byly většinou v souvislosti s jeho ochranný účinek pod biotického stresu na kontrolu preharvest a ztráty po sklizni odvozených z patogenu ovoce infekce (Babalar et al., 2007; Cao et al., 2013). Obecně platí, že endogenní obsah volného SA je vyšší na začátku vývoje ovoce a poté se postupně snižuje (Oikawa et al., 2015). Lu et al. (2015) vykázala sekundární nárůst SA během druhé růstové fáze broskvového ovoce., Exogenní aplikace SA s banány zpoždění procesu zrání se snižuje respirační vzplanutí a dužina se slupkou poměr, stejně jako snižuje aktivitu enzymů souvisejících s buněčnou stěnu degradace a antioxidační systém (Srivastava a Dwivedi, 2000). Kromě toho ošetření sladkých třešní kyselinou salicylovou a kyselinou acetylsalicylovou (ASA) zvyšuje hmotnost, pevnost a barvu třešní v komerční fázi (Giménez et al., 2014). Exogenní aplikace SA nebo metylsalicylátu (MeSA) během po sklizni, zpožděné nadměrné zrání v ovoci, jako jsou kiwi (Zhang et al.,, 2003) a sladké třešně (Valero et al., 2011). Kromě toho ošetření salicylovou nebo ASA zmírnilo mrazivé zranění granátových jablek (Sayyari et al., 2011), rajčata (Aghdam et al., 2014) a avokádo (Glowacz et al., 2017). SA také interaguje s ethylenem inhibicí exprese ACC oxidázy (ACO) (Shi a Zhang, 2012). Překvapivě, zvýšený obsah exprese ACS u hrušek se také nachází po aplikaci SA v dávce regulovaným způsobem (Shi et al., 2013; Shi a Zhang, 2014). Proto je regulace výroby ethylenu závislá na SA jemně regulována (obrázek 2C).,
Melatonin: Vznikající Regulátor pro Řízení Vývoje Ovoce
Melatonin je také rostlinný hormon, z chorismate-odvozené dráhy, který získal velkou pozornost v posledních letech (Hernández-Ruiz et al., 2005; Arnao a Hernández-Ruiz, 2018; Sharif et al., 2018). Nedávno byl melatonin zdůrazněn jako důležitý faktor pro sadu ovoce podobným způsobem jako účinky IAA(obrázek 2a; Liu s. et al., 2018). Kromě toho byla pozorována cirkadiánní produkce tohoto fytohormonu (Kolář et al., 1997)., Během dozrávání na stromě vykazovaly hrozny ošetřené melatoninem vyšší synchronicitu a zvýšenou hmotnost při úplném zrání(Meng et al ., 2015). Navíc další nedávná studie Xu et al. (2018) popsal, že melatonin by mohl podporovat zrání hroznů prostřednictvím interakce s ABA a ethylenem, stejně jako peroxid vodíku. Bylo prokázáno, že melatonin poskytuje chlazení tolerance ovoce během skladování v chladu, které se zdá být většinou souvisí s jeho antioxidační aktivitou (Cao et al., 2018; Aghdam et al., 2019)., Během po sklizni rajčata vykazovala zvýšené emise ethylenu po aplikaci melatoninu na 50 µM (Sun et al., 2015), na rozdíl od hrušek, kde exogenní aplikace melatoninu při 100 µM inhibovala produkci ethylenu (Zhai et al., 2018) nebo banánové ovoce, kde melatonin při 50-200 µM inhiboval expresi ACS a ACO (obrázek 2c; Hu et al., 2017). Ačkoli různé účinky aplikace melatoninu mohou souviset s dávkovacím účinkem, je zapotřebí dalšího výzkumu, aby se lépe pochopila role melatoninu při kontrole vývoje ovoce.,
Cross-Talk Chorismate Odvozené od Hormonů v Řízení Vývoje Ovoce
Hormonální přeslechy mezi chorismate odvozené od hormonů bylo prokázáno, že se vyskytují v průběhu vývoje ovoce a zrání. Breitel et al. (2016) zjistili, že auxiny mohou komunikovat s SA prostřednictvím ARF2 (Obrázek 2C) a že zvýšená exprese ARF2 v rajčatové za následek nižší obsah SA a významné zpoždění zrání., To ukazuje, že auxiny mohou být omezující SA produkci ovoce během vývoje, což by mohlo vysvětlovat, trade-off mezi ovoce růst (zprostředkované auxin) a aktivace biotické obranu (zprostředkované SA). Kromě toho, exogenní aplikace SA na papája ovoce vyústil ve změněné expresi několika IAA geny, některé z nich jsou down-regulovány, zatímco jiní up-regulovaných (Liu et al., 2017)., V tomto případě, výsledky byly neprůkazné, co se opravdu vyskytuje endogenně v domnělé cross-talk mezi auxinu a SA během zrání ovoce a je nutný další výzkum v této a jiných klimakterických plodů. V každém případě je zřejmé, že IAA a SA jsou úzce spjaty nejen metabolicky, ale i funkčně. A konečně, několik studií uvádí vliv melatoninu na IAA a SA biosyntézy a/nebo signalizační (přezkoumána v Arnao a Hernández-Ruiz, 2018), i když je nám známo, žádná z těchto studií byla provedena ani v období menopauzy, ani v non-klimakterický ovoce.,
Modulace Bioaktivních Sloučenin Biosyntézu Chorismate-Odvozené Fytohormony
Bioaktivní sloučeniny (včetně fenolických sloučenin, isoprenoids, a antioxidační vitamíny) nejenže byly široce zkoumány jako zodpovědné za specifické organoleptické vlastnosti potravin, ale také pro své ochranné účinky na lidské buňky před oxidačních procesů, v rozvoji neurodegenerativních a kardiovaskulárních onemocnění a určitých typů rakoviny (Liu et al., 1999; Mueller et al., 2010; Sturgeon a Ronnenberg, 2010; Björkman et al., 2011)., Fenolické sloučeniny významně přispět k předávání specifických chutí, jako jsou třísloviny – zodpovědné za hořkost nebo trpkost chuť některých druhů ovoce, a barvy, jako je anthocyanin – odpovědné za červené, modré a fialové barvy ovoce (Croteau et al., 2000). Význam fenolických sloučenin pro lidskou spotřebu byla spojena s ochranným účinkem proti oxidační procesy ve vztahu ke kardiovaskulární a centrální nervový systém zdraví, stejně jako snižuje riziko rakoviny trávicího traktu (Björkman et al., 2011)., Karotenoidy, tetraterpenes patřící do isoprenoids, hrají roli v ochraně proti foto-oxidačních procesů a jako organické pigmenty; jsou zodpovědné za oranžové-žluté barvy ovoce (Tapiero et al., 2004). Předpokládá se, že dietní karotenoidy poskytují zdravotní přínosy při snižování rizika očních onemocnění a některých typů rakoviny kvůli jejich roli antioxidantů (Johnson, 2002). Vitamíny C A E, včetně kyseliny askorbové a tokoferolů, jsou základními faktory nutriční kvality v ovoci s mnoha biologickými aktivitami u lidí (Miret a Müller, 2017)., S ohledem na příslušné úlohy všech těchto bioaktivních sloučenin jako zdraví prospěšné sloučeniny v ovoci, modulace jejich hromadění je prvořadý význam v obou klimakterium a non-klimakterické plody (Vasconsuelo a Boland, 2007). Většinu těchto bioaktivních látek se hromadí na vysoké úrovni během zrání ovoce, kdy méně chutné zelené plody converse do nutričně bohaté, barevné a chutné ovoce (Daood et al., 1996; Ranalli et al., 1998; Dumas et al., 2003; Singh et al., 2011)., Jakmile však začne, zrání nemůže být zastaveno a obecně vede k nadměrnému zrání, které negativně ovlivňuje kvalitu ovoce (Kumar et al., 2014). Proto minimalizace znehodnocení ovoce po sklizni při získávání vysokého obsahu bioaktivních sloučenin zůstává jednou z největších výzev k vyřešení.
Auxin byl identifikován jako důležitý regulátor biosyntézy karotenoidů během dozrávání klimakterických plodů rajčat. Zrání v rajčatové je spojena s rozkladem chlorofylu a posun xantofyly na karoteny (beta-karoten a lykopen) (Fraser et al., 1994)., IAA se zdá zpoždění rajčat zrání tím, že potlačují ethylenu a několik upstream karotenoid přepisy, včetně Psy, Ziso, Pds, Critiso stejně jako Chlases 1-3 a, na druhé straně, podporovat hromadění β-Lyc1 a Crtr-β1 přepisy, což má za následek vyšší obsah xantofyly a chlorofylu a (Su et al., 2015). Kromě toho, v některých non-klimakterický ovoce, jako jsou třešně a hroznové bobule auxin také se zdá modulovat anthocyanin biosyntézu, a tím kontrolu zrání ovoce procesů. Teribia et al., (2016) vykázala negativní korelaci mezi obsahem IAA a obsahem antokyaninu, což naznačuje, že akumulace antokyaninu začíná, když se obsah IAA snižuje. Navíc, aplikace syntetického auxinu, jako sloučenina benzothiazole-2-oxyacetic kyseliny (BTOA) zpoždění up-regulace genů, které podporují enzymy anthocyanin biosyntéza jako rna syntázy a UDP-glukózy:flavonoid 3-O-glucosyltransferase v hroznové bobule (Davies et al., 1997).,
Pre – a post-sklizni ošetření s salicyláty, včetně SA a její deriváty ASA a MeSA, byly hlášeny regulovat bioaktivních látek akumulace, což vede k lepší antioxidační aktivitu v obou klimakterium a non-klimakterický ovoce. Je známo, že několik druhů ovoce, jako je švestka, třešeň a meruňky, má krátkou životnost s rychlým zhoršením kvality po sklizni. Proto existuje neustálé hledání léčby, které zlepšují a udržují kvalitu ovoce, a zejména obsah bioaktivních sloučenin s příznivými účinky na zdraví., Ošetření švestek SA, ASA a MeSA vedlo k významnému vyššímu obsahu kyseliny askorbové, antokyanů a fenolických sloučenin jak při sklizni, tak po dlouhodobém skladování za studena (40 dní při 4°C; Martínez-Esplá et al., 2017). Podobné výsledky lze pozorovat u léčby preharvest s SA, ASA a MeSA ve sladkých třešních (Giménez et al., 2014, 2015, 2017). Kromě toho, ošetření po sklizni s SA, ASA, nebo MeSA zachován celkový obsah fenolických stejně jako anthocyanin obsah při skladování v granátového jablka (Sayyari et al., 2011), sladká třešeň (Valero et al.,, 2011), cornelian cherry (Dokhanieh et al., 2013) a meruňka (Wang et al., 2015). Tyto výsledky naznačují, že salicyláty mohou být zapojeny do aktivace fenylalaninu amoniak lyázy, což je hlavní enzym podílející se na biosynthesic cesta z fenolické sloučeniny (Martínez-Esplá et al., 2017).
nedávno bylo prokázáno, že melatonin reguluje zrání ovoce a moduluje bioaktivní sloučeniny. Ošetření po sklizni melatoninem v broskvovém ovoci zvýšilo mrazivou toleranci aktivací antioxidačních systémů., Chlazení zranění se vyskytuje v broskev ovoce během skladování při nízké teplotě (mezi 2 a 7,6°C), vyznačující se tím, že maso do zhnědnutí, abnormální zrání a vyšší citlivost k rozpadu (Lurie a Crisosto, 2005). Cao et al. (2018) uvádí, že broskve léčeni melatoninu ukázaly vyšší přepis množství kyseliny askorbové geny biosyntézy PpGME, PpGPP, a PpGLDH na sedmém a PpGMPH na 21. a 28. den skladování ve srovnání s kontrolní což vede ke zvýšení kyseliny askorbové obsah., Navíc melatonin indukovaný zvyšuje aktivity g6pdh, SKDH a PAL, které jsou základními enzymy pro biosyntézu fenolických sloučenin. Autoři naznačují, že léčba melatoninem chrání broskvové ovoce do určité míry před mrazivým zraněním specificky aktivací biosyntézy fenolických sloučenin (Gao et al., 2018). Kromě toho analýza diferenciální proteomiky bez štítků odhalila účinek melatoninu na podporu dozrávání ovoce a akumulaci anthocyaninu po sklizni v rajčatovém ovoci (Sun et al., 2016)., Autoři uvádí, že exogenní melatonin se zvýšil osm enzymy vztahující se k anthocyanin cestou, včetně mimo jiné flavonol-3-hydroxylázy, flavanone 3 beta-hydroxylázy, anthocyanidin syntázy/leucoanthocyanidin dioxygenase, a anthocyanidin 3-O-glucosyltransferase. Kromě toho, ošetření po sklizni s melatonin zvýšení celkové fenoly a antokyany v jahoda ovoce (Aghdam a Fard, 2016), a zpoždění ztrátě celkové fenoly, flavonoidy a antokyanů v liči ovoce (Zhang et al., 2018)., Léčba melatoninem zvýšila obsah fenolů, antokyanů a flavonoidů v hroznových bobulích (Xu et al., 2017). Kromě toho, melatonin bylo hlášeno zvýšení lykopen akumulace a výroba ethylenu v rajčatech, což naznačuje, že melatonin může zvýšit obsah lykopenu tím ovlivňuje biosyntézu ethylenu a signalizace (Sun et al., 2015)., Účast ethylenu v melatonin-indukované nařízení bioaktivních sloučenin byla také pozorována v hroznové bobule, kde double block léčba ethylenu ukázal, snižuje účinky melatoninu na obsah polyfenolů (Xu et al., 2017).
závěr
dospělo se k závěru, že fytohormony odvozené od chorismátu, včetně auxinu, SA a melatoninu, sdílejí nejen společný prekurzor, ale hrají zásadní roli při regulaci růstu a dozrávání ovoce., Metabolický a funkční křížový rozhovor mezi nimi a jinými fytohormony se vyskytuje prostorově, aby se jemně reguloval vývoj klimakterických a ne-klimakterických plodů. Rozdíly v dynamice ve klimakterium a non-klimakterický ovoce důkaz, že reakce chorismate-odvozené hormony, není univerzální, ale spíše silně druhově specifické. Kromě toho fytohormony odvozené od chorismátu také modulují akumulaci bioaktivních sloučenin, čímž ovlivňují kvalitu ovoce., Další výzkum je však potřeba, aby lépe pochopit, jak (i) ostatní hormony, jako je ethylen a ABA, modulovat jejich biosyntéza studium klíčových metabolických rozptýlení bodů, (ii) jsou vzájemně propojeny ve funkční a molekulární úrovni, a (iii), které společně modulují bioaktivních sloučenin, biosyntéza a následně vliv nejen ovoce, růst a zrání, ale také kvalitu klimakterium a non-klimakterický ovoce.
Autor Příspěvky
SM-B koncipován a navržen přezkum s pomocí MP-L. MP-L, PM, MM, SM-B napsal rukopis., MP-L připravené údaje s pomocí PM. Všichni autoři přispěli k diskusi o nápadech, revidovali a schválili konečný rukopis.
financování
tato práce byla financována cenou ICREA Academia udělenou SM-B katalánskou vládou.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Böttcher, C., Boss, P. K., and Davies, C. (2011)., Acyl substrát preference IAA-amido syntetázy účet pro změny v hroznové (Vitis vinifera L.) bobule zrání způsobené různými auxinovými sloučeninami indikující význam auxin konjugace ve vývoji rostlin. J.Exp. Bota. 62, 4267–4280. doi: 10.1093/jxb/err134
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Glowacz, M., Bille, M., Tinyane, P. P., a Sivakumar, D. (2017)., Udržování posklizňové kvalitní studené uloženy ‚Hass‘ avokádo, tím, že změní mastných kyselin, obsah a složení s použitím přírodních těkavých látek – methyl jasmonate a methylsalicylát. J. Potravina. Agricu. 97, 5186–5193. DOI: 10.1002 / jsfa.8400
PubMed Abstract / CrossRef Full Text / Google Scholar
Zeng, W., and He, s. y. (2010). Významnou roli flagellin receptor FLAGELLIN-SENSING2 v mediaci stomatal reakci na Pseudomonas syringae pv tomato DC3000 v Arabidopsis. Rostlinný Fyziol. 153, 1188–1198. doi: 10.1104 / s. 110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















