Biologi for Majors jeg
Læringsresultater
- Beskrive den rolle af RNA-polymerase
Dette afsnit vil udvide på det specifikke rolle RNA-polymeraser i løbet af transskription. Læs videre for at lære rollen som RNA-polymeraser i hvert trin af transkription.,
Indledningen af Transskription
i Modsætning til den prokaryote polymerase, der kan binde til en DNA-skabelon på sin egen, eukaryoter kræver flere andre proteiner, der kaldes transkriptionsfaktorer, at første bind til arrangøren region, og derefter hjælpe med at rekruttere den relevante polymerase.
De Tre eukaryote RNA-polymeraser
funktionerne ved eukaryotisk mRNA-syntese er markant mere komplekse dem for prokaryoter. I stedet for en enkelt polymerase omfattende fem underenheder, eukaryoter har tre polymeraser, der hver består af 10 underenheder eller mere., Hver eukaryotisk polymerase kræver også et særskilt sæt transkriptionsfaktorer for at bringe det til DNA-skabelonen.
RNA-polymerase I er placeret i nucleolus, en specialiseret nuklear understruktur, hvor ribosomalt RNA (rRNA) transkriberes, behandles og samles til ribosomer (tabel 1). RRNA-molekylerne betragtes som strukturelle RNA ‘ er, fordi de har en cellulær rolle, men ikke oversættes til protein. RRNA ‘ erne er komponenter af ribosomet og er afgørende for oversættelsesprocessen. RNA-polymerase i syntetiserer alle rRNA ‘ erne bortset fra 5s rRNA-molekylet., “S” – betegnelsen gælder for” Svedberg ” – enheder, en ikke-additiv værdi, der karakteriserer den hastighed, hvormed en partikelsediment under centrifugering.
| Tabel 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Eukaryote præ-mRNA ‘ er gennemgår omfattende behandling efter transkription, men før oversættelse (Figur 1). For klarhedens skyld vil dette moduls diskussion af transkription og oversættelse i eukaryoter bruge udtrykket “mRNAs” til kun at beskrive de modne, forarbejdede molekyler, der er klar til at blive oversat. RNA-polymerase II er ansvarlig for transkribering af det overvældende flertal af eukaryote gener.

Figur 1. Eukaryotisk mRNA indeholder introner, der skal splejses ud. En 5′ cap og 3 ‘ poly-A hale er også tilføjet.,
RNA-polymerase III er også placeret i kernen. Denne polymerase transkriberer en række strukturelle RNA ‘er, der inkluderer 5S pre-rRNA, transfer pre-RNA’ er (pre-tRNA ‘er) og små nukleare pre-RNA’ er. TRNA ‘ erne har en kritisk rolle i oversættelse; de tjener som adaptermolekylerne mellem mRNA-skabelonen og den voksende polypeptidkæde. Små nukleare RNA ‘ er har en række funktioner, herunder “splejsning” pre-mRNA ‘ er og regulering af transkriptionsfaktorer.,
en videnskabsmand, der karakteriserer et nyt gen, kan bestemme, hvilken polymerase der transkriberer det ved at teste, om genet udtrykkes i nærvær af en bestemt svampegift, a-amanitin (tabel 1). Interessant nok påvirker a-amanitin produceret af Amanita phalloides, Death Cap-svampen, de tre polymeraser meget forskelligt. RNA-polymerase I er fuldstændig ufølsom over for a-amanitin, hvilket betyder, at polymerasen kan transkribere DNA in vitro i nærvær af denne gift. I modsætning hertil er RNA-polymerase II ekstremt følsom over for a-amanitin, og RNA-polymerase III er moderat følsom., At kende den transkriberende polymerase kan knytte en forsker til den generelle funktion af genet, der studeres. Da RNA-polymerase II transkriberer langt de fleste gener, vil vi fokusere på denne polymerase i vores efterfølgende diskussioner om eukaryote transkriptionsfaktorer og promotorer.
RNA-Polymerase II-promotorer og transkriptionsfaktorer
eukaryote promotorer er meget større og mere indviklede end prokaryote promotorer. Imidlertid har begge en sekvens svarende til -10-sekvensen af prokaryoter., I eukaryoter kaldes denne sekvens Tata-boksen og har konsensussekvensen TATAAA på kodningsstrengen. Den er placeret ved -25 til -35 baser i forhold til initieringsstedet (+1) (Figur 2). Denne sekvens er ikke identisk med E. coli -10-boksen, men den bevarer det A–T-rige element. Termostabiliteten af A-T-bindinger er lav, og dette hjælper DNA-skabelonen til lokalt at slappe af som forberedelse til transkription.,i stedet for den enkle factor-faktor, der hjælper med at binde den prokaryote RNA-polymerase til dens promotor, samler eukaryoter et kompleks af transkriptionsfaktorer, der kræves for at rekruttere RNA-polymerase II til et proteinkodende gen. Transkriptionsfaktorer, der binder til promotoren, kaldes basale transkriptionsfaktorer. Disse basale faktorer kaldes alle TFII (til transkriptionsfaktor/polymerase II) plus et yderligere bogstav (A-J). Kernekomplekset er TFIID, som indeholder et Tata-bindende protein (TBP)., De andre transkriptionsfaktorer systematisk at falde på plads på DNA-skabelon, med hver en yderligere stabilisering af pre-indledning komplekse og bidrager til rekruttering af RNA-polymerase II.
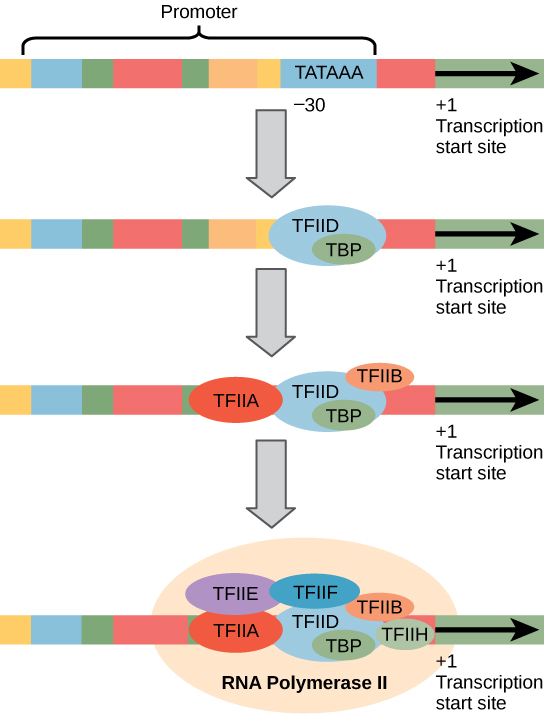
Figur 2. En generaliseret promotor af et gen transkriberet af RNA-polymerase II er vist. Transkriptionsfaktorer genkender promotoren. RNA-polymerase II binder og danner derefter transkriptionsinitieringskomplekset.,
praksis spørgsmål
en videnskabsmand splejser en eukaryotisk promotor foran et bakterielt gen og indsætter genet i et bakterielt kromosom. Ville du forvente, at bakterierne transkriberer genet?
nogle eukaryote promotorer har også en konserveret CAAT-boks (GGCCAATCT) ved cirka -80. Yderligere opstrøms for Tata-boksen kan eukaryote promotorer også indeholde en eller flere GC-rige kasser (GGCG) eller octamer-kasser (ATTTGCAT)., Disse elementer binder cellulære faktorer, der øger effektiviteten af transkriptionsinitiering og identificeres ofte i mere “aktive” gener, som konstant udtrykkes af cellen.
basale transkriptionsfaktorer er afgørende i dannelsen af et præinitieringskompleks på DNA-skabelonen, der efterfølgende rekrutterer RNA-polymerase II til transkriptionsinitiering. Kompleksiteten af eukaryotisk transkription slutter ikke med polymeraser og promotorer., En hær af andre transkriptionsfaktorer, der binder til opstrømsforstærkere og lyddæmpere, hjælper også med at regulere den frekvens, hvormed pre-mRNA syntetiseres fra et gen. Forstærkere og lyddæmpere påvirker effektiviteten af transskription, men er ikke nødvendige for transskription at fortsætte.
udviklingen af promotorer
udviklingen af gener kan være et velkendt koncept. Mutationer kan forekomme i gener under DNA-replikation, og resultatet kan eller ikke være gavnligt for cellen., Ved at ændre et en .ym, strukturelt protein eller en anden faktor kan mutationsprocessen omdanne funktioner eller fysiske egenskaber. Eukaryote promotorer og andre genregulerende sekvenser kan dog også udvikle sig. Overvej for eksempel et gen, der gennem mange generationer bliver mere værdifuldt for cellen. Måske koder genet et strukturelt protein, som cellen skal syntetisere i overflod for en bestemt funktion. Hvis dette er tilfældet, ville det være gavnligt for cellen for genets promotor at rekruttere transkriptionsfaktorer mere effektivt og øge genekspression.,
forskere, der undersøger udviklingen af promotorsekvenser, har rapporteret forskellige resultater. Til dels skyldes det, at det er svært at udlede præcis, hvor en eukaryot promotor begynder og slutter. Nogle promotorer forekommer inden for gener; andre er placeret meget langt opstrøms, eller endda nedstrøms, af de gener, de regulerer. Når forskere begrænsede deres undersøgelse til menneskelige kernepromotorsekvenser, der eksperimentelt blev defineret som sekvenser, der binder preinitiationskomplekset, fandt de imidlertid, at promotorer udvikler sig endnu hurtigere end proteinkodende gener.,
det er stadig uklart, hvordan promotorudvikling kan svare til udviklingen af mennesker eller andre højere organismer. Imidlertid er udviklingen af en promotor til effektivt at fremstille mere eller mindre af et givet genprodukt et spændende alternativ til udviklingen af generne selv.
Promotor Strukturer for RNA Polymeraser i og III
De processer, for at bringe RNA-polymeraser i og III til DNA-skabelon indebære en anelse mindre komplekse samlinger af transkriptionsfaktorer, men det overordnede tema er den samme.,
Den bevares promotor elementer for gener transskriberet af polymeraser i og III adskiller sig fra dem, der er transskriberet af RNA-polymerase II. RNA-polymerase jeg overfører gener, der har to GC-rige promotor-sekvenser i -45 til +20 regionen. Disse sekvenser alene er tilstrækkelige til, at transkriptionsinitiering kan forekomme, men promotorer med yderligere sekvenser i regionen fra -180 til -105 opstrøms for initieringsstedet vil yderligere forbedre initieringen. Gener, der transkriberes af RNA-polymerase III, har opstrøms promotorer eller promotorer, der forekommer i generne selv.,
eukaryotisk transkription er en tæt reguleret proces, der kræver en række proteiner til at interagere med hinanden og med DNA-strengen. Selvom transkriptionsprocessen i eukaryoter indebærer en større metabolisk investering end i prokaryoter, sikrer den, at cellen transkriberer nøjagtigt de præ-mRNA ‘ er, som den har brug for til proteinsyntese.,
Forlængelse og Afslutning
Efter dannelsen af preinitiation komplekse, polymerase er frigivet fra andre transkriptionsfaktorer, og en forlængelse er tilladt at fortsætte, som det gør i prokaryotes med polymerasen syntetiserer præ-mRNA 5′ til 3′ – retningen. Som tidligere omtalt transkriberer RNA-polymerase II den største andel af eukaryote gener, så dette afsnit vil fokusere på, hvordan denne polymerase opnår forlængelse og opsigelse.,
selvom den en .ymatiske forlængelsesproces i det væsentlige er den samme i eukaryoter og prokaryoter, er DNA-skabelonen mere kompleks. Når eukaryote celler ikke deler sig, eksisterer deres gener som en diffus masse af DNA og proteiner kaldet kromatin. DNA ‘ et er tæt pakket omkring ladede histonproteiner med gentagne intervaller. Disse DNA-histonkomplekser, samlet kaldet nukleosomer, er regelmæssigt fordelt og inkluderer 146 nukleotider af DNA, der er viklet omkring otte histoner som tråd omkring en spole.,
for at polynukleotidsyntese kan forekomme, skal transkriptionsmaskineriet flytte histoner ud af vejen, hver gang det støder på et nukleosom. Dette opnås ved et specielt proteinkompleks kaldet FACT, som står for “letter kromatintranskription.”Dette kompleks trækker histoner væk fra DNA-skabelonen, når polymerasen bevæger sig langs den. Når præ-mRNA er syntetiseret, erstatter FAKTAKOMPLEKSET histonerne for at genskabe nukleosomerne.
afslutningen af transkription er forskellig for de forskellige polymeraser., I modsætning til i prokaryoter finder forlængelse af RNA-polymerase II i eukaryoter sted 1.000-2.000 nukleotider ud over slutningen af genet, der transkriberes. Denne pre-mRNA hale fjernes efterfølgende ved spaltning under mRNA-behandling. På den anden side kræver RNA-polymeraser i og III termineringssignaler. Gener transkriberet af RNA-polymerase i indeholder en specifik 18-nukleotidsekvens, der genkendes af et termineringsprotein. Processen med opsigelse i RNA-polymerase III involverer et mRNA-hårnål svarende til rho-uafhængig afslutning af transkription i prokaryoter.,
prøv det
bidrage!
forbedre denne sidelær mere
- H Liang et al., “Hurtig udvikling af kernepromotorer i primatgenomer”, Molekylærbiologi og Evolution 25 (2008): 1239-44. ↵















