Grænser i Plant Science
Indledning
Korrekt progression af frugt modning er væsentlig for at opnå en optimal frugt kvalitet og lang holdbarhed, er vigtige egenskaber, der bestemmer prisen markeder, og endelig overskud. Modning er imidlertid en kompleks proces, hvor mange faktorer er involveret, herunder hormonel kontrol, der regulerer biokemiske og fysiologiske ændringer, der giver endelige organoleptiske og ernæringsmæssige frugtegenskaber., Traditionelt er kødfulde frugter blevet klassificeret i henhold til der modningsproces til klimakteriske og ikke-klimakteriske frugter. Climacteric frugter såsom tomater eller bananer kræver en stigning i respirationshastighed og ethylenproduktion for at frigøre modningsprocessen i et autokatalytisk respons (Barry og Giovannoni, 2007; Liu et al., 2015)., Antagonistically, ikke-climacteric frugter som citrusfrugter og vindruer ikke udviser en burst-hverken i ethylen eller respiration, før modning debut, og i disse frugter, modning er primært kontrolleret af gradvis ophobning af phytohormone abscisinsyre (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Ikke desto mindre forstærker nye undersøgelser ideen om, at modningsprocessen ikke kun styres af produktionen af en phytohormon, men snarere synes at være resultatet af en kontrolleret hormonbalance (Symons et al., 2012; Teribia et al., 2016; Li et al., 2019).,AU auxiner, salicylsyre (SA) og melatonin er fytohormoner involveret i signalering og regulering af mange vigtige processer i planter. Auxiner i vid udstrækning blevet beskrevet som vækst og udvikling regulatorer med flere funktioner i planter (se Taylor-Teeples et al. For 2016); SA udløser forsvar reaktion mod biotrophic og hemi-biotrophic patogener (Loake og Grant, 2007), samt at have en vigtig rolle under abiotisk stress (Dong et al., 2014; Wanani et al., 2017), blomstring og cellecykluskontrol (Cars .ell et al., 1989; Eberhard et al.,, Henholdsvis 1989); og melatonin har ikke kun vist sig at have AU .inlignende funktioner (Chen et al., 2009; 2009uo et al., 2014; 2014en et al. For 2016) men det har også været foreslået at fungere som en potentiel antioxidant i visse planter (Arnao og Hernández-Ruiz, 2015) og en regulator af plantens reaktion på patogener (Chen et al., 2018). Interessant, disse tre hormoner deler en fælles forløber-chorismate -, således en metabolisk cross-talk opstår mellem dem, og en række gener skal være fint reguleret at aflede chorismate metabolisme mod disse forbindelser., I løbet af de sidste par år har flere anmeldelser behandlet AU .ins biosyntese og rolle (Paul et al., 2012), SA (Asghari og Aghdam, 2010) og melatonin (Feng et al., 2014; Arnao og Herndende and-Rui and, 2018) i frugt. Imidlertid er deres biosyntese og funktioner for det meste blevet beskrevet separat. Her vil vi diskutere de fælles og differentielle aspekter af biosyntesen, metabolisme og funktion af Au .in, SA og melatonin i væksten og modningen af climacteric og ikke-climacteric frugter., Hovedvægten vil blive lagt på metaboliske omlægning centrale punkter i deres biosyntese fra chorismate, regulerende rolle i modning af climacteric og ikke-climacteric frugter, og deres rolle i modulation af biosyntesen af bioaktive stoffer, der i vid udstrækning bestemmer, frugt kvalitet.
Chorismatafledte phytohormoner
Chorismat er det endelige produkt af shikimatvejen og spiller en nøglerolle i biosyntesen af phytohormoner (Figur 1)., Chorismate giver anledning til aromatiske aminosyrer, herunder tryptofan, og gennem en række reaktioner, herunder konverteringen af tryptofan til indol-3-pyrodruesyre af tryptophan aminotransferase (TA), og det kan generere auxin indol-3-eddikesyre (IAA). TA er blevet foreslået som en universel nøgle enzym til IAA biosynthesis ikke kun i vegetative organer (Enríquez-Valencia et al., 2018), men også i udviklingen af reproduktive organer (Reyes-Olalde et al., 2017), herunder frugtvækst og modning (Estrada-Johnson et al., 2017)., I modsætning hertil, når isochorismatsyntase (ICS) aktiveres, kan chorismat omdannes til isochorismat, og sidstnævnte kan omdannes til SA. På trods af at denne vej først blev identificeret i bakterier, er den også i øjeblikket veletableret i planter (.ildermuth et al., 2001; Uppalapati et al., 2007; Catinot et al., 2008; Garcion et al., 2008; Abreu og munn and-Bosch, 2009). Endelig kan tryptophan blive tryptamin af en .ymet tryptophan decarbo .ylase (TDC)., Selvom dette er en anden af de flere ruter af IAA-biosyntese (tryptamin bliver til indol-3-acetaldehyd og senere til IAA), er TDC også blevet foreslået at være det første hastighedsbegrænsende en .ym i melatoninvejen (Kang et al., 2007). TDC blev først identificeret i Apocynaceae-familien (De Luca et al., 1989), der senere skal beskrives i en række plantesystemer (Byeon et al., 2012, 2014; 2012hao et al., 2013; 2013ei et al., 2018)., Efter decarboxylation, tryptamin er omdannet til serotonin ved tryptamin-5-hydroxylase, og derefter serotonin omdannes til N-acetyl-serotonin til endelig at give melatonin i cytosol (Figur 1).
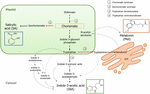
Figur 1. Biosyntesen af indol-3-eddikesyre (IAA), salicylsyre (SA) og melatonin (Mel) fra chorismate. Alle tre forbindelser deler en fælles endelig forløber (chorismate, kvadreret i orange) fra shikimate Path .ay., Større omdirigeringspunkter er angivet med røde pile, og de tilsvarende en .ymer er nummereret i en nedstigningsdivergensrækkefølge fra shikimate. Fed stiplede pile angiver mere end et trin mellem forbindelser. Tynde stiplede pile angiver veje, der stadig ikke er veldefinerede.
Nøgle-omstilling af enzymer, der fører til syntese af chorismate-vej hormoner er for nylig blevet identificeret i kødfulde frugter (Figur 1). ICS, hvilket udtryk har vist sig at være afgørende for biotisk stresstolerance (Garcion et al.,, 2008; 2010eng og He, 2010), er identificeret i tomater og æbler. Zhu et al. (2016) rapporterede OVEREKSPRESSIONEN af ICS1 (isochorismatsyntase 1) under kolde opbevaringsforhold i tomat, mens .hang et al. (2017) fundet aktivering af en transkriptionsfaktor involveret i den patogenrelaterede SA-signalvej, der inducerer ICS-ekspression i æbler. Det er rapporteret, at forbedret TDC-aktivitet forekommer i umodne peberfrugter ved infektion med patogener-gennem øget ekspression af tdc1 og tdc2 (Park et al., 2013 – – og i vækststadiet i morbærfrugt (Wanang et al., 2016)., Endelig er TA identificeret i grapevine, både ved præ-og v .raison (B .ttcher et al., 2013; Gouthu og deluc, henholdsvis 2015) såvel som under jordbær modning (Estrada-Johnson et al., 2017). Det er dog bemærkelsesværdigt, at de gener og enzymer, der er beskrevet i climacteric frugter (dvs, ICS) er ikke blevet identificeret i ikke-climacteric frugter (dvs, TDC og TA) og vice versa, og det er derfor, at yderligere undersøgelser er nødvendige for at udfylde disse huller i den eksisterende viden og til bedre at forstå, hvordan disse omlægning punkter er reguleret i fællesskab i løbet af frugt modning.,
rolle af Chorismatafledte fytohormoner i klimakteriske og ikke-klimakteriske frugter
Unraveling mekanismerne for frugtudvikling har været en af de største udfordringer i nyere agronomiforskning for dens økonomiske konsekvenser. I denne sammenhæng er phytohormoner blevet påpeget som ansvarlige drivkræfter for frugt modning, specielt ethylen og ABA i henholdsvis climacteric og ikke-climacteric frugter. Imidlertid, at disse fytohormoner kunne regulere frugtudvikling alene, viste sig snart at være alt for enkle., Efter omfattende forskning og med forbedringen i Analytisk Kemi og molekylære teknikker er flere andre hormoner blevet bekræftet som potentielle regulatorer af frugtudvikling og modning, herunder chorismatafledte phytohormoner.
Auxiner Cross-Talk Med Andre Hormoner i Løbet af Frugt, der er, Vækst og Modning
Auxiner er en gruppe af plante-hormoner, som spiller en afgørende rolle i frugt udvikling, både at udøve deres egen indflydelse og modulerende udtryk for andre plantehormoner., Endogent indhold af IAA er særligt højt ved frugtsæt og i de indledende vækstudviklingsstadier, hvorefter IAA-mængder har en tendens til at falde inden modning begynder, både i climacteric (.aharah et al., 2012) og ikke-klimakteriske frugter (Symons et al., 2012; Teribia et al., 2016), med tilsyneladende nogle undtagelser, som ferskner (Tatsuki et al., 2013) og nogle blommesorter (El-Sharka .y et al., 2014; figur 2A). Det er påvist, at IAA er involveret i initiering af frugtsæt i kombination med gibberelliner (me..etti et al., 2004; Serrani et al., 2010; Bermejo et al., 2018; Hu et al.,, 2018). Forringelse af IAA-biosyntese eller signalering fører generelt til frugt parthenocarpy, skønt det også kan resultere i unormal modning i nogle frugter (.ang et al., 2005; Liu J. et al., 2018; Reig et al., 2018). Et højt indhold af IAA i de indledende faser af frugt udvikling fremme frugt vækst på grund af auxin konsekvenser i celledeling i kombination med cytokinins og i kontrollen af cellernes ekspansion i kombination med gibberellins (Liao et al., 2018)., I denne periode tillader hormonel krydstale mellem au .iner og gibberelliner desuden normal frugtformning i en finjusteret regulering medieret af Au .in-Responsfaktorer (ARFs; Liao et al., 2018; Liu S. Et al., 2018).
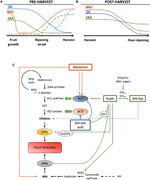
Figur 2. Rolle IAA, SA, og Mel under udviklingen af overgangsalderen og ikke-overgangsalderen frugter. Model, der opsummerer interaktionerne mellem IAA, SA og Mel under modningen af climacteric og ikke-climacteric frugter under (a) før og (B) efter høst., Stiplede linjer angiver alternativ dynamik af fytohormonindhold i nogle frugter (Se tekst til diskussion). C) oversigt over interaktionen mellem IAA, SA og Mel og ethylen og abscisinsyre (ABA) biosyntese i climacteric og ikke-climacteric frugter. Auxin er en positiv regulator af ethylen biosyntesen af aktiveringen af ACC-syntase gener (ACS) og inducere udtryk for Ethylen Lydhør Faktorer (ERFs). Auxin også undertrykker ABA-produktion ved 9-cis-epoxycarotenoid dioxygenase (NCED) undertrykkelse og hæmme ABA-responsive stress gener (ARS) af ARF2 protein., Endogent IAA-indhold kan reduceres med GH3-proteiner, hvis syntese fremmes af ethylen, ABA og sukkerarter. Diskontinuerlig linje angiver IAA ‘ s mulige rolle af IAA – asp som en modningsfaktor, der forbedrer ethylen i nogle frugter. Melatonin forbedrer ABA-og ethylenproduktionen, selvom ACS kan hæmmes afhængigt af melatoninindholdet. SA virker som en hæmmer af ethylen, der undertrykker ACC o .idase (ACO) gener, selv om det kan fremme ekspressionen af ACS., MTA, 50-methylthioadenosine; S-AdoMet, S-adenosyl-methionin; SAM-syntase, S-adenosyl-methionin syntase; ACC, 1-aminocyclopropane-1-carboxylic acid; IPP, isopentenyl diphosphat.
reduktioner af endogent IAA-indhold er rapporteret at forekomme inden modningens begyndelse i flere frugter. Disse reduktioner har været relateret til IAA-konjugering med asparaginsyre (IAA-Asp) af IAA-amido-syntetaser, GH3-1 og GH3-2 (Figur 2C)., Faktisk viste GH3-1 og GH3-2 højere ekspression under tidlig frugtudvikling og især under modningsinitiering både i klimakteriske frugter, såsom tomater (Sravankumar et al., 2018) og æbler (Onik et al ., 2018) såvel som I ikke-klimakteriske frugter, som druebær (B .ttcher et al., 2010, 2011) og hindbær (Bernales et al., 2019)., Interessant nok viste druebær forbedret GH3-1-ekspression efter ABA – og ethephon-applikation, hvilket kunne forklare inddragelsen af ethylen i kontrollen af IAA-indhold efter modningens begyndelse, selv i ikke-klimakteriske frugter (B .ttcher et al., 2010). Faktisk fremhæver flere undersøgelser den stramme interaktion mellem au .iner og ethylen i frugtmodning med en gensidig indflydelse mellem dem (Tadiello et al., 2016a; Busatto et al., 2017)., For climacteric frugter, øget indhold af IAA er nødvendige for at aktivere udtryk for ACC-syntase gener (ACS), kodning ACC-syntase, som igen vil føre til produktion af ethylen, der udløser modningsprocessen (Figur 2C; Tatsuki et al., 2013). Frugter som tomater (Li et al ., 2016) eller bananer (Choudhury et al.,, 2008), som normalt opleve reducerede IAA indholdet forud for påbegyndelsen af modning, viser en modning forsinkelse, når IAA eller-analoger er exogenously anvendt, mens frugter som ferskner, hvis IAA indholdet stige gradvist, indtil det er fuldt modne, vis modning fremskyndet, når auxiner er anvendt (Tadiello et al., 2016b). Derfor skal der findes en stram, kompleks og differentieret regulering af Au .in-ethyleninteraktioner i forskellige frugter under modning før høst., Endvidere IAA indholdet fald før modning indtræden i nogle ikke-climacteric frugter ved at undertrykke 9-cis-epoxycarotenoid dioxygenase (NCED) og starte ABA-syntese (Figur 2C; Jia et al., 2016). Interessant nok formidles denne proces også af IAA-konjugation gennem forbedret GH3-1-aktivitet (B .ttcher et al., 2010). Ikke desto mindre Estrada-Johnson et al. (2017) viste, at selv om IAA indholdet fald i løbet af jordbær modning, udtryk for FaAux/IAA og FaARF gen familier er induceret i røde beholdere, hvilket tyder på inddragelse af auxin signalering i fuldt ud modne frugter.,
Under post-harvest, auxin indholdet normalt forblive uforandret, eller har tendens til at falde på grund af oxidative processer, der kan give anledning til små, men gradvis reduktion i de endogene indhold af IAA (Figur 2B). AU .in behandlinger efter høst forsinkelse over-modning i nogle frugter (Chen et al., 2016; Moro et al., 2017) og øge indholdet af nogle organiske syrer, opretholde frugt surhed (Li et al., 2017), hvilket antyder, at Au .in også spiller en betydelig rolle i kontrollen med frugtmodning under efterhøsten.,
Flere Roller af Salicylsyre I Frugt Udvikling og Modning
Salicylsyre er en anden chorismate-afledte phytohormone, der har været mest knyttet til sin beskyttende effekt under biotisk stress til kontrol preharvest og post-høst tab, der stammer fra patogen frugt infektion (Babalar et al., 2007; Cao et al., 2013). Generelt er endogent indhold af fri SA højere i begyndelsen af frugtudviklingen og falder derefter gradvist (Oika .a et al., 2015). Lu et al. (2015) rapporterede en sekundær stigning i SA i den anden vækstfase af ferskenfrugter., Eksogene anvendelse af SA i bananer forsinket modning proces at mindske den respiratoriske burst og papirmasse til peel-forhold, samt faldende aktivitet af enzymer, der er relateret til cellevæg, nedbrydning og antioxidant system (Srivastava og Dwivedi, 2000). Desuden øgede behandling af søde kirsebærtræer med salicylsyre og acetylsalicylsyre (asa) vægt, fasthed og farve på kirsebær på kommercielt Stadium (Gimnene.et al., 2014). Eksogen anvendelse af SA eller metylsalicylat (Mesa) under Efterhøst, forsinket overmodning i frugter som Ki .ier (methang et al.,, 2003) og søde kirsebær (Valero et al ., 2011). Endvidere lindrede behandlinger med salicylsyre eller ASA køleskade af granatæbler (Sayyari et al., 2011), tomater (Aghdam et al., 2014), og avocadoer (Glo .ac.et al., 2017). SA interagerer også med ethylen gennem hæmning af ACC – o .idase (ACO) – ekspression (Shi and 2012hang, 2012). Overraskende findes øget indhold af ACS-ekspression i pærer også efter SA-anvendelse på en dosisreguleret måde (Shi et al., 2013; Shi og andhang, 2014). Regulering af ethylenproduktion afhængig af SA er derfor fint reguleret (figur 2C).,
Melatonin: En ny Regulator til Styring af Frugt Udvikling
Melatonin er også en plante hormon fra chorismate-afledte vej, der har fået meget opmærksomhed i de seneste år (Hernández-Ruiz et al., 2005; Arnao og Hernández-Ruiz, 2018; Sharif et al., 2018). For nylig er melatonin blevet påpeget som en vigtig faktor for frugtsæt på samme måde som IAA-effekter (figur 2A; Liu S. et al., 2018). Desuden er der observeret cirkadisk produktion af denne fytohormon (Kol.et et al., 1997)., Under modning på træet viste druer behandlet med melatonin højere synkronicitet og øget vægt, når de modnes fuldt ud (Meng et al., 2015). Desuden er en anden nylig undersøgelse af .u et al. (2018) beskrevet, at melatonin kunne fremme drue berry modning gennem sin interaktion med ABA og ethylen, samt hydrogenperoxid. Det er blevet påvist, at melatonin giver køletolerance under frugtkølet opbevaring, hvilket synes at være mest relateret til dets antio .idantaktivitet (Cao et al., 2018; Aghdam et al., 2019)., Under efterhøsten viste tomater øget ethylenemission efter påføring af melatonin ved 50 µM(Sun et al ., 2015), i modsætning til pærer, hvor eksogen anvendelse af melatonin ved 100 µM hæmmede ethylenproduktion (.hai et al., 2018) eller bananfrugter, hvor melatonin ved 50-200 µM hæmmede ekspressionen af ACS og ACO (figur 2C; Hu et al., 2017). Selvom forskellige virkninger af melatoninapplikation kan være relateret til en doseringseffekt, er der behov for yderligere forskning for bedre at forstå melatonins rolle i kontrollen med frugtudvikling.,
Cross-Talk af Chorismatafledte phytohormoner i kontrollen med frugtudvikling
hormonel krydsning mellem chorismatafledte phytohormoner har vist sig at forekomme under frugtudvikling og modning. Breitel et al. (2016) fandt, at auxiner kan interagere med SA gennem ARF2 (Figur 2C), og der overexpression af ARF2 i tomat resulteret i lavere indhold af SA og en væsentlig modning forsinkelse., Dette indikerer, at auinsiner kan begrænse SA-produktionen under frugtudvikling, hvilket kan forklare en afvejning mellem frugtvækst (medieret af Au .in) og aktivering af biotiske forsvar (medieret af SA). Desuden resulterede eksogen anvendelse af SA på papayafrugter i et ændret udtryk for flere IAA-gener, hvoraf nogle blev nedreguleret, mens andre opregulerede (Liu et al., 2017)., I dette tilfælde var resultaterne ufuldstændige af, hvad der virkelig forekommer endogent i den formodede krydssnak mellem au .in og SA under frugtmodning, og der kræves yderligere forskning i denne og andre klimakteriske frugter. Under alle omstændigheder er det klart, at IAA og SA er nært beslægtede, ikke kun metabolisk, men også funktionelt. Endelig er der flere undersøgelser har rapporteret en indflydelse melatonin på IAA og SA biosyntese og/eller signalering (anmeldt i Arnao og Hernández-Ruiz, 2018), selv om det til vores kendskab, at ingen af disse undersøgelser blev udført hverken i climacteric eller i ikke-climacteric frugter.,
Modulation af Bioaktive Stoffer Biosyntesen af Chorismate-Afledte Plantehormoner
Bioaktive stoffer (herunder phenolforbindelser, isoprenoids, og antioxidant-vitaminer) har ikke kun været meget undersøgt som er ansvarlig for særlige organoleptiske egenskaber af fødevarer, men også for deres beskyttende virkninger i menneskets celler mod oxidative processer i udviklingen af neurodegenerative og hjerte-kar-sygdomme og visse typer af kræft (Liu et al., 1999; Mueller et al., 2010; Sturgeon and Ronnenberg, 2010; Bjrrkman et al., 2011)., Phenolforbindelser bidrage væsentligt til at videregive specifikke varianter, såsom tanniner – ansvarlig for bitterhed eller munden smag af visse frugter, og farver, som anthocyanin pigmenter – ansvarlig for rød, blå og lilla frugt farver (Croteau et al., 2000). Relevansen af fenoliske forbindelser til konsum har været forbundet med en beskyttende virkning mod oxidative processer i relation til hjerte-kar-og centralnervesystemet sundhed samt en reduceret risiko for kræft i mave-tarmkanalen (Björkman et al., 2011)., Carotenoider, tetraterpenes, der tilhører isoprenoids, spiller en rolle i beskyttelsen mod foto-oxidative processer, og som organiske pigmenter; de er ansvarlige for de orange-gul farve af frugt (Tapiero et al., 2004). Kosten carotenoider menes at give sundhedsmæssige fordele ved at mindske risikoen for øjensygdom og visse typer kræft på grund af deres rolle som antio .idanter (Johnson, 2002). Vitaminer C og E, herunder henholdsvis ascorbinsyre og tocopheroler, er væsentlige ernæringsmæssige kvalitetsfaktorer i frugter med mange biologiske aktiviteter hos mennesker (Miret and M .ller, 2017)., I betragtning af den relevante rolle af alle disse bioaktive forbindelser som sundhedsfremmende forbindelser i frugter er moduleringen af deres akkumulering af afgørende betydning i både klimakteriske og ikke-klimakteriske frugter (Vasconsuelo og Boland, 2007). De fleste af disse bioaktive forbindelser akkumuleres i høje niveauer under frugtmodning, når mindre velsmagende grønne frugter konverserer til en ernæringsmæssigt rig, farvet og velsmagende frugt (Daood et al., 1996; Ranalli et al., 1998; Dumas et al., 2003; Singh et al. , 2011)., Når modningen først er påbegyndt, kan modningen ikke stoppes og fører generelt til overmodning, der negativt påvirker frugtkvaliteten (Kumar et al ., 2014). Derfor er minimering af frugtforkølelse efter høst, samtidig med at der opnås et højt indhold af bioaktive forbindelser, en af de største udfordringer at løse.
Auxin er blevet identificeret som en vigtig regulator af carotenoid biosynthesis under modning af climacteric tomat frugt. Modning i tomat er forbundet med nedbrydningen af chlorophyll og skiftet af caranthophylls til carotener (β-caroten og lycopen) (Fraser et al., 1994)., IAA ser ud til at forsinke tomat modning ved at undertrykke ethylen og flere opstrøms carotenoid udskrifter herunder Psy, Ziso, Pd, Critiso samt Chlases 1-3, og på den anden side, at fremme ophobning af β-Lyc1 og Crtr-β1 afskrifter, hvilket resulterer i en højere indhold af xantofyler og klorofyl a (Su et al., 2015). Derudover synes AU .in i nogle ikke-klimakteriske frugter, såsom kirsebær-og druebær, også at modulere anthocyaninbiosyntese og derved kontrollere frugtmodningsprocesser. Teribia et al., (2016) rapporterede en negativ korrelation mellem IAA og anthocyaninindhold, hvilket indikerer, at anthocyaninakkumulering starter, når IAA-indholdet falder. Desuden er anvendelsen af syntetiske auxin-lignende stof benzothiazol-2-oxyacetic syre (BTOA) forsinket op-regulering af gener, der fremmer enzymer af anthocyanin biosynthesis som chalcone syntase og UDP-glukose:flavonoid-3-O-glucosyltransferase i druer, bær (Davies et al., 1997).,
Pre – og post-harvest behandlinger med salicylater, herunder SA og dets derivater ASA og MeSA, er blevet rapporteret til at regulere bioaktive stoffer ophobning, der fører til forbedrede antioxidant aktivitet i både climacteric og ikke-climacteric frugter. Flere frugter som blomme, kirsebær og abrikoser er kendt for at have en kort levetid med en hurtig forringelse af kvaliteten efter høst. Derfor er der en konstant søgning efter behandlinger, der forbedrer og opretholder frugtkvaliteten, og især indholdet af bioaktive forbindelser med gavnlige sundhedseffekter., Behandlinger af blommer med SA, Asa og MeSA resulterede i betydeligt højere indhold af ascorbinsyre, anthocyaniner og phenolforbindelser både ved høst og efter langvarig køleopbevaring (40 dage ved 4.C; Mart .ne.-Espl. et al., 2017). Lignende resultater kunne observeres ved præharvest-behandlinger med SA, Asa og MeSA i søde kirsebær (Gim .ne.et al., 2014, 2015, 2017). Endvidere opretholdt behandling efter høst med SA, ASA eller MeSA det samlede phenolindhold såvel som anthocyaninindhold under køleopbevaring i granatæble (Sayyari et al., 2011), sød kirsebær (Valero et al.,, 2011), cornelian cherry (Dokhanieh et al., 2013), og abrikos (Wanang et al ., 2015). Disse resultater antyder, at salicylater kan være involveret i aktiveringen af phenylalanin ammoniaklyase, som er det vigtigste en .ym involveret i den biosyntetiske vej af phenolforbindelser (Mart .ne.-Espl. et al., 2017).for nylig har melatonin vist sig at regulere frugt modning og modulere bioaktive forbindelser. Efter høst behandlinger med melatonin i fersken frugt øget afkøling tolerance ved at aktivere antioxidant-systemer., Nedkøling skade opstår i fersken frugt under opbevaring ved lav temperatur (mellem 2 og 7.6°C) præget af kød bruning, unormal modning og højere følsomhed over for forfald (Lurie og Crisosto, 2005). Cao et al. (2018) rapporterede, at ferskner, der er behandlet med melatonin viste en højere udskrift overflod af ascorbinsyre biosyntese gener PpGME, PpGPP, og PpGLDH på syvende og PpGMPH på 21st og 28 dages opbevaring i forhold til kontrol, hvilket resulterer i øget ascorbinsyre indhold., Desuden inducerede melatonin stigninger i aktiviteter af G6PDH, SKDH og PAL, som er de essentielle en .ymer for phenolforbindelser biosyntese. Forfatterne antyder, at melatoninbehandling beskytter ferskenfrugt til en vis grad mod køleskade ved specifikt at aktivere biosyntesen af phenolforbindelser (Gao et al., 2018). Derudover afslørede en etiketfri differentiel proteomikanalyse effekten af melatonin på at fremme frugtmodning og anthocyaninakkumulering efter høst i tomatfrugter (Sun et al., 2016)., Forfatterne rapporterede, at eksogene melatonin er steget otte enzymer, relateret til anthocyanin vej, herunder bl.a. flavonol-3-hydroxylase, flavanone 3 beta-hydroxylase, anthocyanidin syntase/leucoanthocyanidin dioxygenase, og anthocyanidin 3-O-glucosyltransferase. Desuden, efter høst, behandling med melatonin stigning i de samlede phenoler og anthocyaniner i jordbær frugt (Aghdam og Fard, 2016), og forsinket tab af total phenoler, flavonoider og anthocyaniner i litchi frugt (Zhang et al., 2018)., Behandlinger med melatonin øgede indholdet af phenoler, anthocyaniner og flavonoider i druebær (Melu et al ., 2017). Derudover er det rapporteret, at melatonin forbedrer lycopenakkumulering og ethylenproduktion i tomater, hvilket antyder, at melatonin kan øge indholdet af lycopen ved at påvirke ethylenbiosyntese og signalering (Sun et al., 2015)., Ethylens deltagelse i den melatonininducerede regulering af bioaktive forbindelser er også observeret i druebær, hvor dobbeltblokbehandling af ethylen viste reducerede virkninger af melatonin på polyphenolindhold (.u et al., 2017).
Konklusion
Det er konkluderet, at chorismate-afledte plantehormoner, herunder auxin, SA og melatonin, der ikke kun deler en fælles forløber, men spiller vigtige roller i reguleringen af frugt vækst og modning., En metabolisk og funktionel cross-talk mellem dem og med andre phytohormoner forekommer på en spatiotemporal måde for fint at regulere udviklingen af climacteric og ikke-climacteric frugter. Forskellene i dynamikken inden for klimakteriske og ikke-klimakteriske frugter viser, at responsen af chorismatafledte hormoner ikke er universel, men snarere stærkt artsspecifik. Endvidere modulerer chorismatafledte fytohormoner også akkumuleringen af bioaktive forbindelser, hvilket påvirker frugtkvaliteten., Yderligere forskning er dog nødvendigt for at bedre at forstå, hvordan (jeg) andre hormoner, såsom ethylen og ABA, modulere deres biosyntese studere vigtige metaboliske omlægning point, (ii) de interagerer på det funktionelle og molekylære niveauer, og (iii) de i fællesskab modulere bioaktive stoffer biosyntese og dermed indflydelse ikke kun frugt vækst og modning, men også kvaliteten af climacteric og ikke-climacteric frugter.
Forfatterbidrag
SM-b udtænkt og designet anmeldelsen ved hjælp af MP-L. MP-L, PM, MM og SM-b skrev manuskriptet., MP-L forberedte figurer ved hjælp af PM. Alle forfattere bidrog til diskussionen af ideer, revideret og godkendt det endelige manuskript.
finansiering
dette arbejde blev finansieret af en Icrea Academia a .ard givet til sm-b af den catalanske regering.
Erklæring om interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
b .ttcher, C., Boss, P. K., and Davies, C. (2011)., Acyl substrat præferencer af en IAA-amido synthetase højde for variationer i drue (Vitis vinifera L.) berry modning som følge af forskellige auxinic forbindelser, der angiver betydningen af auxin konjugation i planternes udvikling. J. E Expp. Bot. 62, 4267–4280. doi: 10.1093/jxb/err134
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., og Sivakumar, D. (2017)., Opretholdelse af efterhøstkvaliteten af koldlagrede Hass-avocadoer ved at ændre fedtsyreindholdet og sammensætningen ved brug af naturlige flygtige forbindelser – methyl jasmonat og methylsalicylat. J. Sci. Fødevare. Agric. 97, 5186–5193. doi: 10.1002/jsfa.8400
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
Zeng, W., og Han, S. Y. (2010). En fremtrædende rolle af flagellinreceptoren FLAGELLIN-SENSING2 i formidlende stomatal respons på Pseudomonas syringae pv tomat DC3000 i Arabidopsis. Plantefysiol. 153, 1188–1198. doi: 10.1104 / s.110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















