Biologian suuret Yhtiöt en
osaamistavoitteet
- Kuvaile rooli RNA-polymeraasi
Tämä osio laajenee, kun erityinen rooli RNA-polymerases aikana transkriptio. Lue oppia rooli RNA polymeraasien kussakin vaiheessa transkription.,
Aloittamisesta Transkriptio
toisin Kuin prokaryooteilla-polymeraasi, joka voi sitoutua DNA: n mallin omasta, eukaryooteissa vaatia useita muita proteiineja kutsutaan transkriptio tekijät, ensin sitoa promoottori alueen ja sitten auttaa rekrytoimaan sopiva polymeraasi.
Kolme Aitotumallisilla RNA-Polymerases
ominaisuudet eukaryoottisesta mRNA synteesi ovat selvästi monimutkaisempia kuin prokaryootit. Viiden alayksikön muodostaman yhden polymeraasin sijaan eukaryooteilla on kolme polymeraasia, jotka kukin koostuvat 10 alayksiköstä tai useammasta., Jokainen aitotumallisilla-polymeraasi vaatii myös selkeitä transkriptio tekijät tuoda sen DNA-malli.
RNA-polymeraasi I sijaitsee vuonna nucleolus, erikoistunut ydinvoiman alusrakenne jossa ribosomaalisen RNA (rRNA) on puhtaaksi, käsitellään ja kootaan ribosomit (Taulukko 1). Se rRNA molekyylit pidetään rakenteellisia RNAs, koska ne ovat solujen rooli, mutta niitä ei ole käännetty proteiini. Rrnat ovat ribosomien komponentteja ja välttämättömiä käännösprosessille. RNA polymeraasi I syntetisoi kaikki rrnat lukuun ottamatta 5S rRNA-molekyyliä., ”S” – nimitys koskee ”Svedberg” yksikköä, nonadditive arvo, joka luonnehtii nopeus, jolla hiukkasta sedimenttien sentrifugoinnin aikana.
| Taulukko 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Eukaryoottiset pre-mrnat läpikäyvät laajan käsittelyn transkription jälkeen, mutta ennen käännöstä (Kuva 1). Selvyyden vuoksi todetaan, että tämä moduuli on keskustelua, transkriptio ja käännös eukaryooteissa käyttää termiä ”mRNAs” kuvaamaan vain kypsä, käsitellä molekyylejä, jotka ovat valmiita käännetty. RNA polymeraasi II vastaa valtaosan eukaryoottisista geeneistä transponoinnista.

Kuva 1. Eukaryoottinen mRNA sisältää introneita, jotka on yhdistettävä pois. Lisäksi lisätään 5′ korkki ja 3 ’ poly-a pyrstö.,
RNA-polymeraasi III sijaitsee myös tumassa. Tämä polymeraasi transcribes erilaisia rakenteellisia RNAs, joka sisältää 5S pre-rRNA -, siirto-pre-RNAs (pre-tRNAs), ja pieni ydin pre-RNAs. Trnat ovat ratkaisevassa asemassa kääntämisessä; ne toimivat mRNA-mallin ja kasvavan polypeptidiketjun muuntajamolekyyleinä. Pieni ydinalan RNAs on erilaisia toimintoja, kuten ”liitos” pre-mRNAs ja säännellä transkriptio tekijät.,
tutkija luonteenomaiset uusi geeni voi määrittää, mitkä polymeraasi transcribes se testaamalla, onko geeni on ilmaistu läsnäollessa erityisesti sieni myrkkyä, α-amanitin (Taulukko 1). Mielenkiintoista, α-amanitin valmistettu kavalakärpässieni, Kuolema Korkki sieni, vaikuttaa kolme polymeraasien hyvin eri tavalla. RNA-polymeraasi I on täysin tunteeton α-amanitin, mikä tarkoittaa, että polymeraasi voi puhtaaksi DNA: n in vitro-läsnäollessa myrkkyä. Sen sijaan, RNA-polymeraasi II on erittäin herkkä α-amanitin, ja RNA-polymeraasi III on kohtalaisen herkkä., Transcribing-polymeraasin tunteminen voi johtaa tutkijan tutkittavan geenin yleiseen toimintaan. Koska RNA-polymeraasi II transcribes valtaosa geenit, me keskitymme tämän polymeraasi meidän myöhemmät keskustelut eukaryoottisista transkriptio tekijät ja toteuttajat.
RNA-Polymeraasi II Vetäjät ja Transkriptio Tekijät
Aitotumallisilla vetäjät ovat paljon suurempia ja enemmän monimutkainen kuin prokaryooteilla vetäjät. Kummassakin on kuitenkin prokaryoottien -10-sekvenssiä muistuttava sekvenssi., Eukaryooteissa tätä sekvenssiä kutsutaan TATA-laatikoksi, ja sillä on konsensusjärjestys TATAAA koodausjuurella. Se sijaitsee pisteessä -25–35 suhteessa aloituspaikkaan (+1) (kuva 2). Tämä sekvenssi ei ole sama kuin E. coli–10 laatikko, mutta se säästää A-T rikas Elementti. Se thermostability At joukkovelkakirjat on alhainen ja tämä auttaa DNA-mallin paikallisesti rentoutua valmisteltaessa transkriptio.,
sen Sijaan, että yksinkertainen σ tekijä, joka auttaa sitoa prokaryoottiset RNA-polymeraasi sen promoottori, eukaryooteissa koota monimutkainen transkriptio tekijät tarvitaan rekrytoida RNA-polymeraasi II proteiinia koodaavan geenin. Transkriptiotekijät, jotka sitoutuvat promoottori kutsutaan pohjapinta transkriptio tekijät. Nämä pohjapinta tekijät ovat kaikki nimeltään TFII (Transkription Tekijä/ – polymeraasi II) plus ylimääräinen kirjain (A-J). Ydinkompleksi on TFIID, johon kuuluu TATA-sitovaa proteiinia (TBP)., Muut transkriptiotekijät järjestelmällisesti loksahtaa kohdalleen DNA: n malli, jossa jokainen edelleen vakauttaa ennen aloittamista monimutkainen ja edistää rekrytointia RNA-polymeraasi II.
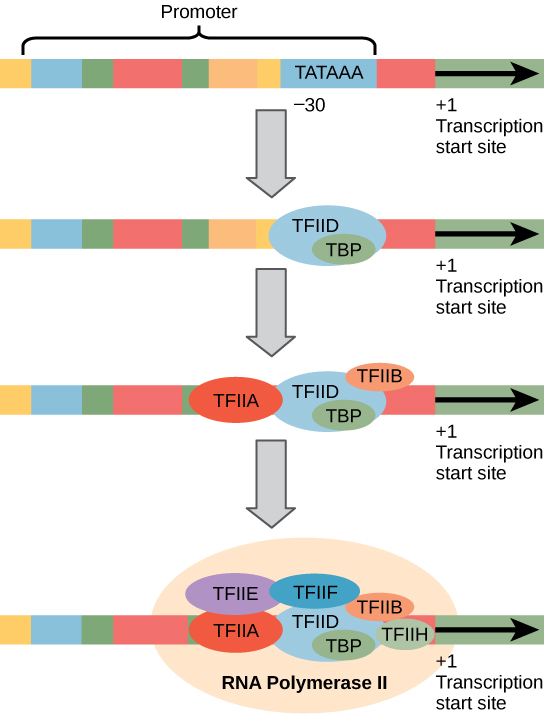
Kuva 2. RNA polymeraasi II: n transkriboiman geenin yleistynyt promoottori on esitetty. Transkriptiotekijät tunnistavat promoottorin. RNA-polymeraasi II sitoo ja muodostaa transkription initiaatiokompleksin.,
Käytännössä Kysymys
tutkija liitosta on aitotumallisilla promoottori edessä bakteeri geeni ja lisää geeni bakteeri kromosomi. Odottaisitko bakteerien siirtävän geenin?
Jotkut aitotumallisilla toteuttajien on myös säilytetty CAAT-laatikko (GGCCAATCT) noin -80. TATA-laatikon yläjuoksulla eukaryoottiset promoottorit voivat sisältää myös yhden tai useamman GC-rikkaan rasian (GGCG) tai oktameerilaatikon (ATTGCAT)., Nämä tekijät sitovat cellular tekijöitä, jotka lisäävät tehokkuutta transkriptio aloittamista, ja ne ovat usein tunnistettu enemmän ”aktiivinen” geenit, jotka ovat jatkuvasti ilmaisseet solun.
Pohjapinta transkriptio tekijät ovat tärkeitä muodostumista preinitiation monimutkainen DNA-malli, joka myöhemmin rekrytoi RNA-polymeraasi II transkriptio aloittamista. Eukaryoottisen transkription monimutkaisuus ei pääty polymeraasiin ja promoottoreihin., Armeija muut transkriptiotekijät, jotka sitoutuvat alkupään parantajia ja äänenvaimentimet, myös auttaa säätelemään taajuus, jolla pre-mRNA: ta syntetisoidaan geeni. Tehostajat ja äänenvaimentimet vaikuttavat transkription tehokkuuteen, mutta ne eivät ole tarpeen transkription jatkamiseksi.
Kehitystä Toteuttajat
kehitys geenejä voi olla tuttu käsite. Geeneissä voi esiintyä mutaatioita DNA: n replikaation aikana, ja tuloksesta voi olla solulle hyötyä tai ei., Muuttamalla entsyymiä, rakenneproteiinia tai jotain muuta tekijää mutaatioprosessi voi muuttaa toimintoja tai fyysisiä ominaisuuksia. Eukaryoottiset promoottorit ja muut geenien säätelysekvenssit voivat kuitenkin myös kehittyä. Ajatellaanpa esimerkiksi geeniä, joka monien sukupolvien aikana muuttuu solulle arvokkaammaksi. Ehkä geeni koodaa rakenteellista proteiinia, jota solun täytyy syntetisoida runsaasti tiettyä tehtävää varten. Jos näin on, se olisi hyödyllistä solun kyseisen geenin promoottori rekrytoida transkriptio tekijät tehokkaammin ja lisätä geenien ilmentyminen.,
promoottorijaksojen evoluutiota tutkivat tutkijat ovat raportoineet vaihtelevia tuloksia. Osittain tämä johtuu siitä, että on vaikea päätellä juuri kun aitotumallisilla promoottori alkaa ja päättyy. Jotkut promoottorit esiintyvät geenien sisällä; toiset sijaitsevat hyvin kaukana ylävirtaan tai jopa alavirtaan säätelemistään geeneistä. Kuitenkin, kun tutkijat rajoitettu niiden tutkiminen ihmisen core promoter sekvenssit, jotka olivat määritelty kokeellisesti, kuten sekvenssit, jotka sitovat preinitiation monimutkainen, he huomasivat, että vetäjät kehittyä jopa nopeammin kuin proteiini koodaus geenien.,
on vielä epäselvää, miten promoottorin evoluutio voisi vastata ihmisten tai muiden korkeampien organismien evoluutiota. Kuitenkin kehitys promoottori tehokkaasti tehdä enemmän tai vähemmän tietyn geenin tuote on kiehtova vaihtoehto evoluutio geenien itse.
Promoottori Rakenteet RNA-Polymerases, I ja III
prosessit tuo RNA-polymerases, I ja III DNA-mallin mukaan hieman vähemmän monimutkaisia kokoelmia transkriptio tekijät, mutta yleinen teema on sama.,
säilytetään promoottori elementtejä geenit transkriptoidaan polymeraasien I ja III eroavat transkriptoidaan RNA-polymeraasi II. RNA-polymeraasi I transcribes geenejä, jotka ovat kaksi GC-rikas promoottori sekvenssit -45 +20 alueella. Nämä sekvenssit eivät yksin riitä transkriptio aloittamista esiintyä, mutta vetäjiä ja muita sekvenssejä alueella -180 että -105 ennen aloittamista sivusto on edelleen parantaa aloittamista. RNA polymeraasi III: n transkriboimissa geeneissä on ylävirtaan eteneviä promoottoreita tai promoottoreita, jotka esiintyvät itse geeneissä.,
Aitotumallisilla transkriptio on tiukasti säädelty prosessi, joka vaatii erilaisia proteiineja vuorovaikutuksessa toistensa kanssa ja DNA-strand. Vaikka prosessi transkriptio vuonna eukaryooteissa liittyy suurempi metabolinen investointi kuin prokaryootit, se takaa sen, että solujen muoto juuri pre-mRNAs, että se tarvitsee proteiinisynteesiä.,
Venymä ja Irtisanominen
Seuraavat muodostumista preinitiation monimutkainen, polymerase vapautuu muut transkriptiotekijät, ja venymä ei saa edetä, koska se ei prokaryootit kanssa-polymeraasi syntetisoi pre-mRNA: n 5′ 3′ suuntaan. Kuten aiemmin keskusteltiin, RNA polymeraasi II transcribes suurin osa eukaryoottisten geenien, joten tässä osassa keskitytään siihen, miten tämä polymeraasi saavuttaa venymä ja päättyminen.,
vaikka venymän entsymaattinen prosessi on eukaryooteissa ja prokaryooteissa olennaisesti sama, DNA-malli on monimutkaisempi. Kun soluille ei jakamalla, niiden geenit ovat olemassa, koska hajanainen massa DNA: ta ja proteiineja kutsutaan chromatin. DNA pakataan tiiviisti latautuneiden histoniproteiinien ympärille toistuvin väliajoin. Nämä DNA–histoni komplekseja, yhdessä kutsutaan jaa twitteriinjaa, ovat säännöllisesti välein ja sisältää 146 nukleotidien DNA haavan noin kahdeksan histones, kuten lanka ympärillä kela.,
polynukleotidin synteesissä transkriptiokoneen on siirrettävä histonit pois tieltä aina, kun se kohtaa nukleosomin. Tämä tapahtuu erityisellä PROTEIINIKOMPLEKSILLA nimeltä fakta, joka tarkoittaa ” helpottaa kromatiinin transkriptiota.”Tämä kompleksi vetää histoneja pois DNA-mallista, kun polymeraasi liikkuu sitä pitkin. Kun pre-mRNA on syntetisoitu, SE monimutkainen korvaa histones luoda jaa twitteriinjaa.
transkription päättyminen on eri polymeraasilla erilainen., Toisin kuin prokaryootit, venymä RNA-polymeraasi II eukaryooteissa tapahtuu 1,000–2,000 nukleotidien päättymisen geeni on puhtaaksi. Tämä pre-mRNA-pyrstö poistetaan myöhemmin pilkkomalla mRNA-käsittelyn aikana. Toisaalta RNA-polymeraasit I ja III vaativat irtisanomissignaaleja. Geenit transkriptoidaan RNA-polymeraasi I sisältävät tietyn 18-sekvenssin, joka on tunnustettu irtisanominen proteiinia. Prosessin päättymisestä RNA-polymeraasi III liittyy mRNA hiusneula samanlainen rho-riippumaton irtisanominen transkriptio vuonna prokaryootit.,
Kokeile
Edistää!
Parantaa tätä pageLearn Enemmän
- S Liang et al., ”Core promoottoreiden nopea kehitys kädellisten genomeissa”, Molecular Biology and Evolution 25 (2008): 1239-44. ↵















