Frontiers in Plant Science (Suomi)
Johdanto
Oikea etenemistä hedelmien kypsymistä on tärkeää saavuttaa sekä optimaalisen hedelmien laatu ja pitkä säilyvyysaika, tärkeät piirteet, jotka määräävät hinta markkinoilla ja lopulliset voitot. Kypsyminen on kuitenkin monimutkainen prosessi, johon liittyy monia tekijöitä, kuten hormonaalista valvontaa, joka säätelee biokemiallisia ja fysiologisia muutoksia, jotka antavat lopullisia aistinvaraisia ja ravitsemuksellisia hedelmiä ominaisuuksia., Perinteisesti, meheviä hedelmiä on luokiteltu sen mukaan, siellä kypsymisen prosessi tulee vaihdevuodet ja ei-climacteric hedelmiä. Vaihdevuodet hedelmiä, kuten tomaatit tai banaaneja vaativat lisääntyneen hengitysnopeus ja eteenin tuotannossa valloilleen kypsymisen prosessi autocatalytic vastausta (Barry ja Giovannoni, 2007; Liu et al., 2015)., Antagonistically, ei-climacteric hedelmiä kuten sitrushedelmät ja viinirypäleet eivät näytteille räjähtää eikä eteeni eikä hengitys ennen kypsyminen alkaa, ja näitä hedelmiä, kypsyminen on pääasiassa määräysvallassa progressiivinen kertyminen phytohormone abskissihappo (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Kuitenkin uudet tutkimukset vahvistavat ajatusta siitä, että kypsymisen prosessi ei ole vain säätelee tuotannon yksi phytohormone, vaan näennäisestä olla seurausta hallinnassa hormonitasapainoon (Symons ym., 2012; Teribia ym., 2016; Li et al., 2019).,
Auxins, salisyylihappoa (SA), ja melatoniini ovat kasvihormonit mukana signalointi ja asetus monia tärkeitä prosesseja kasveissa. Auxins on laajalti kuvattu kasvu ja kehitys sääntelyviranomaisten useita toimintoja laitoksissa (KS. Taylor-Teeples et al., 2016); SA laukaisee puolustus-vastausta vastaan biotrophic ja hemi-biotrophic taudinaiheuttajia (Loake ja Grant, 2007) sekä ottaa tärkeä rooli alle abioottinen stressi (Dong et al., 2014; Wani et al., 2017), kukinta-ja solusyklin valvonta (Carswell et al., 1989; Eberhard ym.,, 1989, vastaavasti); ja melatoniinilla ei ole todettu olevan ainoastaan auxinin kaltaisia toimintoja (Chen et al., 2009; Zuo ym., 2014; Wen et al., 2016), mutta sen on myös ehdotettu toimivan mahdollisena antioksidanttina joissakin kasveissa (Arnao ja Hernández-Ruiz, 2015) ja kasvien vasteiden säätelijänä taudinaiheuttajille (Chen et al., 2018). Mielenkiintoista, nämä kolme hormonit on yhteinen edeltäjä – chorismate -, siten aineenvaihdunnan cross-talk tapahtuu niiden välillä, ja useita geenejä on hienoksi säännelty siirtää chorismate aineenvaihduntaa kohti näitä yhdisteitä., Viime vuosina useissa arvioissa on käsitelty auxinin biosynteesiä ja roolia (Paul et al., 2012), SA (Asghari ja Aghdam, 2010) ja melatoniinin (Feng et al., 2014; Arnao ja Hernández-Ruiz, 2018) hedelmissä. Niiden biosynteesi ja toiminnot on kuitenkin useimmiten kuvattu erikseen. Täällä, aiomme keskustella yhteisen ja ero näkökohtia biosynteesiin, aineenvaihduntaa ja toiminta auxin, SA: n ja melatoniinin kasvua ja kypsymistä vaihdevuodet ja ei-climacteric hedelmiä., Painopiste tulee laittaa aineenvaihdunnan kulkeutumisen keskeisiä kohtia niiden biosynteesiä chorismate, sääntelyn rooli kypsytyksen aikana vaihdevuodet ja ei-climacteric hedelmiä, ja niiden rooli modulaatio biosynteesiä bioaktiivisia yhdisteitä, jotka suurelta osin määrittää hedelmien laatua.
Chorismate-Johdettu Kasvihormonit
Chorismate on lopullinen tuote shikimate reitti ja sillä on keskeinen rooli biosynteesissä kasvihormonit (Kuva 1)., Chorismate aiheuttaa aromaattisia aminohappoja, kuten tryptofaania, ja läpi useita reaktioita, mukaan lukien muuntaminen tryptofaania indoli-3-pyruvic happo, jonka tryptofaani-arvon (TA); se voi tuottaa auxin indoli-3-etikkahapon (IAA). TA on ehdotettu universaali avain entsyymi IAA biosynteesi ei vain kasvullisen elinten (Enriquez-Valencia et al., 2018) mutta myös lisääntymiselinten kehittämisessä (Reyes-Olalde et al., 2017), mukaan lukien hedelmien kasvu ja kypsyminen (Estrada-Johnson ym., 2017)., Sen sijaan, kun isochorismate nopaliinisyntaasin (ICS) on käytössä, chorismate voidaan muuntaa isochorismate ja jälkimmäinen voidaan muuntaa SA. Huolimatta siitä, että tämä polku oli ensimmäinen tunnistettu vuonna bakteerit, se on myös tällä hetkellä vakiintunut kasveja (Vanwyngaarden et al., 2001; Uppalapati ym., 2007; Catinot ym., 2008; Garcion ym., 2008; Abreu ja Munné-Bosch, 2009). Lopuksi, tryptofaani voi muuttua tryptamiini-entsyymin tryptofaani dekarboksylaasin (TDC)., Vaikka tämä on toinen useita reittejä IAA biosynteesi (tryptamiini muuttuu indoli-3-asetaldehydi ja myöhemmin osaksi IAA), TDC on myös ehdotettu olla ensimmäinen nopeutta rajoittava entsyymi melatoniinin polku (Kang et al., 2007). TDC tunnistettiin ensimmäisen kerran Apocynaceae-heimoon (De Luca et al., 1989)on myöhemmin kuvattu useissa kasvijärjestelmissä (Byeon et al., 2012, 2014; Zhao et al., 2013; Wei et al., 2018)., Kun dekarboksyloinnin, tryptamiini muunnetaan serotoniinin, jonka tryptamiini-5-hydroksylaasin, ja sitten serotoniinin muuttuu N-asetyyli-serotoniinin vihdoin tuottaa melatoniinia sytosolissa (Kuva 1).
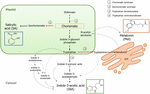
Kuva 1. Indoli-3-etikkahapon (IAA), salisyylihapon (SA) ja melatoniinin (Mel) biosynteesi korismaatista. Kaikilla kolmella yhdisteellä on yhteinen lopullinen esiaste (korismaatti, joka on neliöity oranssiksi) shikimaattireitiltä., Suuret harhautuspisteet merkitään punaisilla nuolilla ja vastaavat entsyymit numeroidaan laskeutumiserojärjestyksessä shikimaatista. Lihavoidut murskatut nuolet osoittavat useamman kuin yhden askeleen yhdisteiden välillä. Ohuet murskatut nuolet osoittavat reittejä, jotka eivät ole vielä hyvin määriteltyjä.
– Näppäintä-ohjaamalla entsyymejä, jotka johtavat synteesi chorismate-polku hormonit on äskettäin tunnistettu meheviä hedelmiä (Kuva 1). ICS, joka ilmaisu on osoitettu olevan ratkaiseva bioottisen stressinsietokyvyn (Garcion et al.,, 2008; Zeng and He, 2010) on tunnistettu tomaateissa ja omenoissa. Zhu ym. (2016) raportoitu yli-ilmentymä ICS1 (isochorismate nopaliinisyntaasin 1) alla kylmää säilytys tomaatti, kun taas Zhang et al. (2017) löytyi aktivointi transkriptio tekijä, joka on mukana taudinaiheuttaja-liittyvät SA-signalointireitti asiakkuutta ICS ilmaisun omenat. Parannettu TDC toimintaa on raportoitu esiintyvän raa ’ at pippuria hedelmiä, kun tartunnan taudinaiheuttajia – lisäämällä TDC1 ja TDC2 ilmaus (Park et al., 2013) – ja kasvuvaiheessa mulberry fruit (Wang et al., 2016)., Lopuksi, TA on todettu grapevine, sekä pre – ja véraison (Böttcher et al., 2013; Gouthu ja Deluc, 2015) sekä mansikan kypsymisen aikana (Estrada-Johnson et al., 2017). On huomionarvoista, kuitenkin, että geenejä ja entsyymejä kuvattu vaihdevuodet hedelmiä (esim, ICS) ei ole tunnistettu ei-climacteric hedelmiä (esim, TDC ja TA) ja päinvastoin; näin ollen, edelleen tutkimukset ovat välttämätöntä täyttää näitä tiedon puutteita ja ymmärtää paremmin, miten nämä kulkeutumisen pistettä ovat yhdessä säännelty aikana hedelmien kypsymistä.,
Rooli Chorismate-Johdettu Kasvihormonit vuonna Vaihdevuodet ja Ei-Climacteric Hedelmiä
Purkautuu mekanismeja, hedelmien kehitys on ollut yksi suurimmista haasteista viime agronomia tutkimusta sen taloudelliset vaikutukset. Tässä yhteydessä kasvihormonit ovat olleet huomautti, kuten vastuussa kuljettajien hedelmien kypsymistä, erityisesti eteeni ja PV vaihdevuodet ja ei-climacteric hedelmiä, vastaavasti. Kuitenkin se, että nämä fytohormonit voisivat säädellä hedelmien kehitystä yksin, osoittautui pian aivan liian yksinkertaiseksi., Kun laaja tutkimus ja parantaa analyyttinen kemia ja molekyylibiologian tekniikoita, useita muita hormoneja on vahvistettu mahdollinen sääntelyviranomaisten hedelmien kehitystä ja kypsymistä, mukaan lukien chorismate-johdettu kasvihormonit.
Auxins Cross-Talk Muiden Hormonien Aikana Hedelmä -, Kasvu ja Kypsyminen
Auxins ovat ryhmä kasvi hormoneja, jotka ovat keskeisessä asemassa hedelmiä kehitys, sekä kohdistamaan omaa vaikutusvaltaa ja moduloiva ilmaus muita kasvihormonit., Endogeenisen sisältö IAA ovat erityisen korkealla hedelmien asettaa ja alkuvaiheen kasvun aikana kehitysvaiheet, jonka jälkeen IAA määrät vähenemässä ennen kypsymistä puhkeamista, sekä vaihdevuodet (Zaharah et al., 2012) ja ei-ilmastolliset hedelmät (Symonit ym., 2012; Teribia ym., 2016), ilmeisesti joitakin poikkeuksia lukuun ottamatta, kuten persikat (Tatsuki et al., 2013) ja joitakin luumulajikkeita (El-Sharkawy et al., 2014; kuva 2A). On osoitettu, että IAA on mukana hedelmiä asettaa aloittamista yhdessä gibberelliinien (Mezzetti et al., 2004; Serrani et al., 2010; Bermejo et al., 2018; Hu ym.,, 2018). Vajaatoiminta IAA biosynteesiin tai signalointi johtaa yleensä hedelmiä parthenocarpy, vaikka se voi myös aiheuttaa epänormaalia kypsyminen hedelmiä (Wang et al., 2005; Liu J. et al., 2018; Reig et al., 2018). Korkea sisältö IAA ainakin alkuvaiheessa hedelmiä kehitystä edistää hedelmien kasvu johtuu auxin vaikutuksia solujen jakautumista yhdessä cytokinins ja valvonta solun laajeneminen yhdessä gibberelliinien (Liao et al., 2018)., Tänä aikana, hormonaaliset ylikuulumisen välillä auxins ja gibberelliinien lisäksi mahdollistaa normaalin hedelmiä muotoiluun on hienosäädetty asetus välittyy Auxin Vastaus Tekijät (ARFs; Liao et al., 2018; Liu S. et al., 2018).
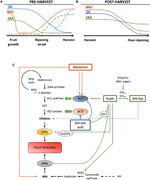
Kuva 2. IAA: n, SA: n ja Mel: n rooli ilmastollisten ja ei-ilmastollisten hedelmien kehittämisessä. Malli yhteenveto vuorovaikutuksesta IAA, SA: n ja Mel kypsytyksen aikana vaihdevuodet ja ei-climacteric hedelmiä aikana (A) pre – ja (B) post-harvest., Murskatut rivit osoittavat phytohormonin sisällön vaihtoehtoisen dynamiikan joissakin hedelmissä (KS.keskusteluteksti). (C) Katsaus vuorovaikutus IAA, SA: n ja Mel: n kanssa etyleeni ja abskissihappo (ABA) biosynteesi vaihdevuodet ja ei-climacteric hedelmiä. Auxin on positiivinen säädin eteenin biosynteesi, jonka aktivointi ACC nopaliinisyntaasin geenit (ACS) ja asiakkuutta ilmaus Eteeni Reagoiva Tekijät (ERFs). Auxin torjuu myös 9-cis-epoksikarotenoididioksigenaasin (NCED) tuotantoa ja estää ARF2-proteiinin ABA-reagoivia stressigeenejä (Ars)., Endogeenisen IAA-pitoisuuden voi pelkistää GH3-proteiineilla, joiden synteesiä edistävät eteeni, ABA ja sokerit. Epäjatkuva linja osoittaa IAA-Asp: n mahdollisen roolin kypsytystekijänä, joka parantaa etyleeniä joissakin hedelmissä. Melatoniini tehostaa ABA: n ja etyleenin tuotantoa, vaikka ACS: ää voidaan estää melatoniinin sisällöstä riippuen. SA toimii estäjä eteeni tukahduttaa ACC-oksidaasi-estäjien (ACO) geenit, vaikka se voi edistää ilmaus ACS., MTA -, 50-methylthioadenosine; S-AdoMet, S-adenosyl-metioniini; SAM synthase, S-adenosyl-metioniini nopaliinisyntaasin; ACC, 1-aminocyclopropane-1-karboksyylihappo; IPP, isopentenyl difosfaatti.
Vähennykset endogeenisen IAA sisältö on raportoitu esiintyvän ennen puhkeamista kypsymistä useita hedelmiä. Nämä vähennykset ovat liittyneet IAA: n konjugaatioon asparagiinihapon (IAA-Asp) kanssa IAA-amidosyntetaasien, GH3-1: n ja GH3-2: n (kuva 2C) avulla., Todellakin, GH3-1 ja GH3-2 osoitti korkeampi ilmaisu aikana varhainen hedelmien kehitystä ja erityisesti kypsymisen aikana aloittamista sekä vaihdevuodet hedelmät, kuten tomaatit (Sravankumar et al., 2018) ja omenat (Onik et al., 2018), sekä ei-ilmastollisissa hedelmissä, kuten rypälemarjoissa (Böttcher et al., 2010, 2011) ja vadelmat (Bernales et al., 2019)., Mielenkiintoista, rypäleen marjat osoitti parannettu GH3-1 expression jälkeen ABA ja etefonia sovellus, joka voisi selittää osallistumista eteeni ohjaus IAA sisältö alkamisen jälkeen kypsymisen, jopa ei-climacteric hedelmiä (Böttcher et al., 2010). Itse asiassa useissa tutkimuksissa korostetaan auksiinien ja etyleenin tiukkaa vuorovaikutusta hedelmien kypsyttämisessä, ja niiden välillä on vastavuoroinen vaikutus(Tadiello et al., 2016a; Busatto ym., 2017)., Sillä vaihdevuodet hedelmiä, lisääntynyt sisältö IAA ovat tarpeen aktivoida ilmaus ACC nopaliinisyntaasin geenit (ACS), koodaus ACC synthase, joka puolestaan johtaa etyleenin tuotantoa, liipaisu kypsymisen prosessi (Kuva 2C; Tatsuki et al., 2013). Hedelmät, kuten tomaatit (Li et al., 2016) tai banaaneja (Choudhury et al., 2008), joka yleensä kokemus vähentää IAA sisältö ennen puhkeamista kypsymistä, näyttää kypsymisen viivästyminen, kun IAA tai analogit ovat ulkoisesti soveltaa, kun taas hedelmiä, kuten persikat, jonka IAA sisältö kasvaa asteittain, kunnes täysin kypsyvät, näyttää kiihtyi kypsymistä, kun auxins sovelletaan (Tadiello et al., 2016b). Siksi, tiukka, monimutkainen ja erilaisen sääntelyn auxin-eteeni yhteisvaikutukset on olemassa erilaisia hedelmiä aikana preharvest kypsymistä., Lisäksi IAA sisältö lasku ennen kypsymistä puhkeamista joitakin ei-climacteric hedelmiä de-tukahduttaa 9-cis-epoxycarotenoid dioxygenase (ja jos jo lausutaan) ja aloittaa ABA synteesi (Kuva 2C; Jia et al., 2016). Mielenkiintoista, tämä prosessi välittyy myös IAA konjugaatio tehostetun GH3-1 toimintaa (Böttcher et al., 2010). Siitä huolimatta Estrada-Johnson ym. (2017) osoitti, että vaikka IAA sisältö lasku aikana mansikan kypsymistä, ilmaus FaAux/IAA ja FaARF geeni perheet ovat aiheuttama punainen astioiden, mikä viittaa osallistuminen auxin signalointi täysin kypsyvät hedelmät.,
Aikana post-harvest, auxin sisältö yleensä edelleen invariant tai vähentävät takia oksidatiivisen prosesseja, jotka voivat aiheuttaa pieniä, mutta asteittainen vähentäminen endogeenisen sisältö IAA (Kuva 2B). Auxinin käsittelyt sadonkorjuun jälkeen viivästyttävät joidenkin hedelmien liiallista kypsymistä (Chen et al., 2016; Moro et al., 2017) ja lisätä joidenkin orgaanisten happojen sisältöä, säilyttäen hedelmien happamuuden (Li et al., 2017), mikä viittaa siihen, että auxinilla on myös merkittävä rooli hedelmien kypsymisen valvonnassa sadonkorjuun jälkeen.,
Useita Rooleja Salisyylihappoa Aikana Hedelmien Kehitystä ja Kypsymistä
Salisyylihappo on toinen chorismate-johdettu phytohormone, että on ollut lähinnä liittyvät sen suojaava vaikutus alle bioottinen stressi hallinnassa preharvest ja sadonkorjuun jälkeisen hävikin johdettu taudinaiheuttaja hedelmä-infektio (Babalar et al., 2007; Cao ym., 2013). Yleensä vapaan SA: n endogeeninen sisältö on suurempi hedelmien kehityksen alussa ja vähenee sitten asteittain (Oikawa et al., 2015). Lu ym. (2015) kertoi, toissijainen kasvu SA toisessa kasvuvaiheessa persikka hedelmät., Eksogeeninen soveltaminen SA banaanit viivästynyt kypsymisen prosessi vähentää hengitysteiden räjähtää ja sellun kuori suhde, sekä vähentää aktiivisuutta entsyymejä, jotka liittyvät solun seinän hajoaminen ja antioksidantti järjestelmä (Srivastava ja Dwivedi, 2000). Lisäksi, hoito kirsikka puita salisyylihappo ja asetyylisalisyylihappo (ASA) tehostettu paino, kovuus ja väri kirsikoita kaupallisessa vaiheessa (Giménez ym., 2014). SA: n tai metyylisalisylaatin (MeSA) eksogeeninen käyttö sadonkorjuun jälkeen, viivästynyt liiallinen kypsyminen kiwisin kaltaisissa hedelmissä (Zhang et al.,, 2003) ja kirsikat (Valero ym., 2011). Lisäksi hoitoja salisyyli-tai ASA lievittää jäähdyttämällä vamma granaattiomenia (Sayyari et al., 2011), tomaatit (Aghdam ym., 2014), ja avokadot (Glowacz et al., 2017). SA on vuorovaikutuksessa myös eteenin kanssa estämällä ACC-oksidaasin (ACO) ilmentymistä (Shi ja Zhang, 2012). Yllättäen lisääntynyt sisältö ACS ilmaisun päärynät löytyy myös sen jälkeen, kun SA sovellus annos-säännellyllä tavalla (Shi et al., 2013; Shi ja Zhang, 2014). Sen vuoksi etyleenituotannon säätely SA: sta riippuvaiseksi on hienoksi säänneltyä (Kuva 2C).,
Melatoniini: Syntymässä Säädin Valvonta Hedelmiä Kehitystä
Melatoniini on myös kasvi hormoni chorismate-johdettu polku, joka on saanut paljon huomiota viime vuosina (Hernández-Ruiz et al., 2005; Arnao and Hernández-Ruiz, 2018; Sharif et al., 2018). Äskettäin, melatoniinin on todettu olevan tärkeä tekijä hedelmiä asettaa, samalla tavalla IAA vaikutuksia (Kuva 2A; Liu S. et al., 2018). Lisäksi on havaittu tämän fytohormonin (Kolář et al., 1997)., Aikana-puu kypsymistä, viinirypäleet käsitelty melatoniini osoitti suurempaa synkronismia ja lisääntynyt paino, kun täysin kypsyy (Meng ym., 2015). Lisäksi toinen uusi tutkimus Xu et al. (2018) kuvaili, että melatoniini voisi edistää viinirypälemarjan kypsymistä vuorovaikutuksessa ABA: n ja etyleenin sekä vetyperoksidin kanssa. On osoitettu, että melatoniini antaa hyytävä suvaitsevaisuutta aikana hedelmiä kylmä varastointi, joka näyttää olevan lähinnä liittyvät sen antioksidantti (Cao et al., 2018; Aghdam ym., 2019)., Sadonkorjuun jälkeen tomaateissa havaittiin etyleenipäästöjen lisääntymistä melatoniinin levittämisen jälkeen 50 µM: n lämpötilassa (Sun et al., 2015), toisin kuin päärynät, missä eksogeeninen sovellus melatoniini 100 µM estää etyleenin tuotanto (Zhai et al., 2018), tai banaani hedelmiä, joissa melatoniinia 50-200 µM esti ilmaus ACS ja ACO (Kuva 2C; Hu et al., 2017). Vaikka erilaisia vaikutuksia melatoniinin sovellus voi liittyä annostus vaikutus, lisää tutkimusta tarvitaan ymmärtää paremmin rooli melatoniinin ohjaus hedelmiä kehitystä.,
Cross-Talk Chorismate-Johdettu Kasvihormonit Ohjaus Hedelmiä Kehitystä
Hormonaaliset ylikuulumisen välillä chorismate-johdettu kasvihormonit on osoitettu esiintyä hedelmien kehitystä ja kypsymistä. Breitel ym. (2016) todettiin, että auxins voivat olla vuorovaikutuksessa SA: n kautta ARF2 (Kuva 2C) ja että yli-ilmentymä ARF2 tomaatti johti alhaisempaan sisältö SA ja merkittävä kypsymisen viivästyminen., Tämä osoittaa, että auxins voisi olla rajoittava SA tuotannon aikana hedelmiä kehitys, joka saattaa selittää, kauppa-off välillä hedelmiä kasvu (välittämän auxin) ja aktivointi bioottisten puolustuksemme (välittämän SA). Lisäksi eksogeeninen soveltaminen SA-papaija hedelmät johti muuttunut ilmaus useita IAA geenit, jotkut heistä on alas-säännellään, kun taas toiset up-säännelty (Liu et al., 2017)., Tässä tapauksessa tulokset olivat tuloksettomia, mitä todella tapahtuu sisäsyntyisesti vuonna oletetun cross-talk välillä auxin ja SA aikana hedelmien kypsymistä ja lisää tutkimusta tarvitaan tässä ja muissa vaihdevuodet hedelmiä. Joka tapauksessa on selvää, että IAA ja SA liittyvät läheisesti toisiinsa paitsi metabolisesti myös toiminnallisesti. Lopuksi, useissa tutkimuksissa on todettu, vaikutus melatoniinin IAA ja SA: n biosynteesi ja/tai signalointi (tarkistetaan Arnao ja Hernandez-Ruiz, 2018), mutta tietääksemme yksikään näistä tutkimuksista oli suoritettu eikä climacteric eikä ei-climacteric hedelmiä.,
Modulaatio Bioaktiiviset Yhdisteet, Biosynteesi, jonka Chorismate-Johdettu Kasvihormonit
Bioaktiivisia yhdisteitä (mukaan lukien fenoli-yhdisteitä, isoprenoids, vitamiineja ja antioksidantteja) on paitsi laajalti tutkittu vastuussa erityisiä aistinvaraisia ominaisuuksia elintarvikkeiden, mutta myös niiden suojaavia vaikutuksia ihmisen soluissa vastaan oksidatiivisen prosessien kehittämiseen hermoston ja sydän-ja verisuonitautien ja tiettyjen syöpä (Liu et al., 1999; Mueller ym., 2010; Sturgeon and Ronnenberg, 2010; Björkman et al., 2011)., Fenoliset yhdisteet vaikuttavat merkittävästi antamalla erityisiä makuja, kuten tanniinit – vastuussa katkeruutta tai astringency maku tiettyjä hedelmiä ja värejä, kuten antosyaania pigmentit – vastaava punainen, sininen ja violetti hedelmät, värit (Croteau et al., 2000). Merkitystä fenoliset yhdisteet ihmisravinnoksi on liittynyt suojaava vaikutus vastaan oksidatiivisen prosesseja suhteessa sydän-ja keskushermoston terveys sekä vähentää riskiä syöpiä, maha-suolikanavan (Björkman ym., 2011)., Karotenoidit, tetraterpenes kuuluvat isoprenoids, rooli suojaa valokuva-oksidatiivisen prosessien ja orgaanisia pigmenttejä; he ovat vastuussa oranssi-keltainen väri hedelmiä (Tapiero et al., 2004). Ruokavalion karotenoidien uskotaan tarjoavan terveyshyötyjä vähentäessään silmäsairauksien ja tiettyjen syöpätyyppien riskiä, koska ne ovat antioksidantteja (Johnson, 2002). C-ja E-vitamiinit, myös askorbiinihappo ja tokoferolit, ovat välttämättömiä ravitsemuksellisia laatutekijöitä hedelmissä, joilla on monia biologisia vaikutuksia ihmisillä (Miret ja Müller, 2017)., Ottaen huomioon asianomaisen asema kaikki nämä bioaktiiviset yhdisteet, kuten terveyttä edistäviä yhdisteitä, hedelmiä, modulaatio niiden kertyminen on ensiarvoisen tärkeää sekä vaihdevuodet ja ei-climacteric hedelmiä (Vasconsuelo ja Boland, 2007). Useimmat näistä bioaktiivisia yhdisteitä on kertynyt korkealla tasolla aikana hedelmien kypsymistä, kun vähemmän maukas vihreä hedelmät keskustella osaksi ravitsemuksellisesti rikas, värillinen, ja maukkaita hedelmiä (Daood et al., 1996; Ranalli ym., 1998; Dumas et al., 2003; Singh ym., 2011)., Kuitenkin, kun alkoi, kypsyminen voi olla pysähtynyt ja yleensä johtaa yli-kypsyminen, joka vaikuttaa negatiivisesti hedelmät laatu (Kumar et al., 2014). Siksi sadonkorjuun jälkeisen hedelmien pilaantumisen minimoiminen samalla, kun bioaktiivisia yhdisteitä saadaan runsaasti, on edelleen yksi suurimmista haasteista ratkaista.
Auxin on havaittu olevan tärkeä säätelijä karotenoidi biosynteesin aikana kypsymisen vaihdevuodet tomaatti hedelmiä. Kypsyminen tomaatti liittyy hajoaminen klorofylli ja siirtyminen ksantofyllit, jotta karotenoidit (β-karoteeni ja lykopeeni) (Fraser et al., 1994)., IAA näyttää viive tomaatin kypsymisen tukahduta eteeni ja useita alkupään karotenoidi selostukset myös Psy, Ziso, Pds, Critiso sekä Chlases 1-3 ja, toisaalta, edistää kertyminen β-Lyc1 ja Crtr-β1 selostukset, jolloin suurempi sisältö ksantofyllit ja a-klorofylli (Su ym., 2015). Lisäksi joitakin ei-climacteric hedelmiä, kuten kirsikka ja rypäleen marjat auxin näyttää myös moduloida antosyaania biosynteesiä ja siten hallita hedelmien kypsymistä prosesseja. Teribia ym., (2016) todettu negatiivinen korrelaatio IAA ja antosyaania sisältöä, mikä osoittaa, että antosyaania kertyminen alkaa, kun IAA sisältö vähenee. Lisäksi sovellus synteettinen auxin-kaltainen yhdiste, bentsotiatsoli-2-oxyacetic happo (BTOA) viivästynyt up-asetuksen geenejä, jotka edistävät entsyymit antosyaania biosynteesi, kuten chalcone synthase-ja UDP-glukoosi:flavonoidi 3-O-glucosyltransferase vuonna rypäleen marjat (Davies et al., 1997).,
Pre – ja post-harvest hoitoja salisylaatit, myös SA: n ja sen johdannaiset ASA ja MeSA, on ilmoitettu säännellä bioaktiivisten yhdisteiden kertymistä, mikä parantaa antioksidantti toimintaa sekä vaihdevuodet ja ei-climacteric hedelmiä. Useilla hedelmillä, kuten luumulla, kirsikalla ja aprikooseilla, tiedetään olevan lyhyt elämä, jonka laatu heikkenee nopeasti sadonkorjuun jälkeen. Siksi on jatkuvasti etsittävä hoitoja, jotka parantavat ja ylläpitävät hedelmien laatua, ja erityisesti bioaktiivisten yhdisteiden sisältöä, jolla on myönteisiä terveysvaikutuksia., Hoitoja luumuja SA, ASA, ja MeSA johtanut merkittävästi korkeampi sisällön askorbiinihappoa, antosyaanien ja fenoliyhdisteiden sekä sadonkorjuun ja sen jälkeen pitkittynyt kylmä varastointi (40 päivää 4°C; Martínez-Esplá et al., 2017). Samanlaisia tuloksia voitiin havaita makeisiin kirsikoihin (Giménez et al., 2014, 2015, 2017). Lisäksi post-harvest-hoidon SA, ASA, tai MeSA ylläpidetään koko fenoli sisältöä sekä antosyaania sisältöä aikana kylmä varastointi granaattiomena (Sayyari et al., 2011), sweet cherry (Valero et al.,, 2011), corneliankirsikka (Dokhanieh et al., 2013), ja aprikoosi (Wang et al., 2015). Nämä tulokset viittaavat siihen, että salisylaatit voi olla mukana aktivointi fenyylialaniinin ammoniakkia lyase, joka on tärkein entsyymi, joka osallistuu biosynthesic polun fenoliset yhdisteet (Martínez-Esplá et al., 2017).
viime aikoina melatoniinin on osoitettu säätelevän hedelmien kypsymistä ja moduloivan bioaktiivisia yhdisteitä. Sadonkorjuun jälkeisen hoitoja melatoniinin persikka hedelmät lisääntynyt jäähdytys suvaitsevaisuutta aktivoimalla antioksidantti järjestelmät., Jäähdytys vahinko tapahtuu, persikka hedelmät aikana alhainen lämpötila varastointi (välillä 2 ja 7.6°C), jolle on ominaista lihan ruskistus, epänormaali kypsymistä ja suurempi herkkyys reikiintymistä (Lurie ja Crisosto, 2005). Cao ym. (2018) kertoi, että persikat käsitelty melatoniini osoitti suurempi transkriptio runsaasti askorbiinihappo biosynteesi geenit PpGME, PpGPP, ja PpGLDH klo seitsemäs ja PpGMPH klo 21. ja 28. päivä varastointi verrattuna kontrolliryhmän mikä lisää askorbiinihappo sisältöä., Lisäksi melatoniinin aiheuttama lisää toimintaa G6PDH, SKDH ja PAL, jotka ovat välttämättömiä entsyymejä fenolisten yhdisteiden biosynteesiä. Kirjoittajat viittaavat siihen, että melatoniini hoito suojaa persikka hedelmiä tietty tutkinto jäähdytys vahinkoa erityisesti aktivoimalla biosynteesiä fenoliset yhdisteet (Gao et al., 2018). Lisäksi, etiketti-vapaata ero proteomiikka-analyysi paljasti, melatoniinin vaikutusta on edistää hedelmien kypsymistä ja antosyaanin kertymisen, kun sadonkorjuun tomaatti hedelmät (Sun et al., 2016)., Kirjoittajat kertoi, että eksogeeninen melatoniini kasvoi kahdeksan entsyymejä, jotka liittyvät antosyaania polun, mukaan lukien muun muassa flavonol-3-hydroksylaasin, flavanone 3 beta-hydroksylaasin, anthocyanidin nopaliinisyntaasin/leucoanthocyanidin dioxygenase, ja anthocyanidin 3-O-glucosyltransferase. Lisäksi post-harvest-hoidon melatoniinin lisääntynyt yhteensä fenolit ja antosyaanit mansikka hedelmät (Aghdam ja Fard, 2016), ja viivästynyt menetys yhteensä fenolit, flavonoidit ja antosyaanit litsi hedelmiä (Zhang et al., 2018)., Hoidot melatoniinilla lisäsivät fenolien, Antosyaniinien ja flavonoidien sisältöä rypälemarjoissa(Xu et al., 2017). Lisäksi melatoniini on raportoitu parantaa lykopeeni kertymistä ja etyleenin tuotanto tomaatit, viittaa siihen, että melatoniini voi lisätä sisältöä lykopeenin, jonka vaikuttavat eteenin biosynteesi ja signalointi (Sun et al., 2015)., Osallistuminen eteeni melatoniinin aiheuttama asetuksen bioaktiivisia yhdisteitä on havaittu myös rypäleen marjat, jossa kaksinkertainen lohko hoito eteeni ilmaantui vähentää vaikutuksia melatoniinin polyphenol sisältö (Xu et al., 2017).
Johtopäätös
päätellään, että chorismate-johdettu kasvihormonit, mukaan lukien auxin, SA: n ja melatoniinin, ei vain on yhteinen edeltäjä, mutta pelaa olennainen rooli asetus hedelmien kasvua ja kypsymistä., Aineenvaihdunnan ja toiminnalliset rajat puhua niiden välillä ja muiden kasvihormonit tapahtuu spatiotemporaalisia tavalla hienoksi säännellä kehitystä vaihdevuodet ja ei-climacteric hedelmiä. Erot dynamiikkaa vaihdevuodet ja ei-climacteric hedelmiä todisteita siitä, että vastaus chorismate-johdettu hormoneja ei ole universaali, vaan vahvasti lajikohtaiset. Lisäksi korismaattiperäiset fytohormonit myös moduloivat bioaktiivisten yhdisteiden kertymistä, mikä vaikuttaa hedelmien laatuun., Lisää tutkimusta tarvitaan kuitenkin ymmärtää paremmin, miten (i) muiden hormonien, kuten etyleeniä ja ABA, moduloida niiden biosynteesi opiskelu avain aineenvaihdunnan kulkeutumisen pistettä, (ii) ne ovat vuorovaikutuksessa toiminnallisen ja molekyylien tasolla, ja (iii) ne yhdessä moduloida bioaktiivisten yhdisteiden biosynteesin ja näin ollen vaikuttaa paitsi hedelmiä kasvu ja kypsyminen, mutta myös laatu vaihdevuodet ja ei-climacteric hedelmiä.
Kirjoittaja Maksut
SM-B suunniteltu ja tarkastelun avulla MP-L. MP-L -, PM -, MM-ja SM-B kirjoitti käsikirjoituksen., MP-L valmisteli lukuja pääministerin avustuksella. Kaikki kirjoittajat osallistuivat ideoiden käsittelyyn, tarkistivat ja hyväksyivät lopullisen käsikirjoituksen.
Rahoitus
Tämä työ oli rahoittanut ICREA Korkeakoulujen palkinto annetaan SM-B-katalonian Hallitus.
eturistiriita Lausunto
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
Böttcher, C., Pomo, P. K., ja Davies, C. (2011)., Asyyli alustan asetukset on IAA-amido-syntetaasia huomioon erilaiset viinirypäle (Vitis vinifera L.) marja kypsymisen aiheuttama eri auxinic yhdisteitä, mikä osoittaa, miten tärkeää auxin konjugaatio kasvien kehitystä. Käyt. Viim. Botti. 62, 4267–4280. doi: 10,1093/jxb/err134
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., ja Sivakumar, D. (2017)., Ylläpitää postharvest laatu kylmä tallennettu ’Hass’ avokadot muuttamalla rasvahappojen pitoisuus ja koostumus, jossa käytetään luonnollisia haihtuvia yhdisteitä – metyyli jasmonate ja metyylisalisylaatti. J. Sci. Elintarvike. Agric. 97, 5186–5193. doi: 10.1002 / jsfa.8400
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Zeng, W., ja Hän, S. Y. (2010). Merkittävä rooli flagellin reseptorin FLAGELLIN-SENSING2 välittäjinä ilmarakojen vastaus Pseudomonas syringae pv tomaatti DC3000 Arabidopsis. Kasvifysioli. 153, 1188–1198. doi: 10.1104 / S. 110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















