Biológia I
tanulási eredmények
- írja le az RNS polimeráz
Ez a szakasz az RNS polimerázoknak a transzkripció során betöltött sajátos szerepére fog kiterjedni. Olvassa tovább, hogy megtanulja az RNS polimerázok szerepét a transzkripció minden szakaszában.,
Megindítása Átírás
Ellentétben a prokarióta polimeráz amely kötődik a DNS-sablon a saját, eukaryotes igényel több más fehérjék, úgynevezett transzkripciós faktorok, hogy első kötődik a promóter régióban majd segíteni toborozni a megfelelő polimeráz.
A három eukarióta RNS polimeráz
az eukarióta mRNS szintézis jellemzői jelentősen összetettebbek a prokarióták jellemzőivel. Az öt alegységből álló egyetlen polimeráz helyett az eukarióták három polimerázzal rendelkeznek, amelyek mindegyike legalább 10 alegységből áll., Minden eukarióta polimeráz külön transzkripciós faktorokat is igényel, hogy a DNS-sablonhoz jusson.
az RNS polimeráz i a nukleolusban található, egy speciális nukleáris szubsztrátumban, amelyben a riboszomális RNS-t (rRNS) átírják, feldolgozzák és riboszómákká alakítják (1.táblázat). Az rRNA molekulákat szerkezeti RNS-eknek tekintik, mivel celluláris szerepük van, de nem fordítják fehérjére. Az rRNAs a riboszóma alkotóelemei, amelyek elengedhetetlenek a fordítási folyamathoz. Az RNS polimeráz I szintetizálja az összes rRNS-t, kivéve az 5S rRNS molekulát., Az” S “jelölés a” Svedberg ” egységekre vonatkozik, egy nemadditív érték, amely jellemzi azt a sebességet, amelyen a részecske üledékek a centrifugálás során.
| 1.táblázat.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Az eukarióta pre-mRNS-ek kiterjedt feldolgozáson mennek keresztül a transzkripció után, de a fordítás előtt (1.ábra). Az egyértelműség kedvéért, ez a modul vita transzkripció és fordítás eukariótákban fogja használni a “mRNAs”, hogy leírja csak az érett, feldolgozott molekulák, amelyek készek lefordítani. Az RNS polimeráz II felelős az eukarióta gének túlnyomó többségének átírásáért.

1.ábra. Az eukarióta mRNS intronokat tartalmaz, amelyeket ki kell illeszteni. Egy 5′-es kupak és 3 ‘ – as poli-a farok is hozzá van adva.,
az RNS polimeráz III szintén a magban található. Ez a polimeráz transcribes a különböző strukturális Rns, amely magában foglalja az 5S előre rrns, átadása pre-Rns (pre-tRNAs), valamint a kis nukleáris pre-Rns. A trnáknak kritikus szerepük van a fordításban; ezek az mRNS sablon és a növekvő polipeptidlánc közötti adaptermolekulák. A kis nukleáris RNS-ek számos funkcióval rendelkeznek, beleértve az mRNS előtti “splicing”-et és a transzkripciós tényezők szabályozását.,
egy új gént jellemző tudós meghatározhatja, hogy melyik polimeráz átírja azt annak tesztelésével, hogy a gént egy adott gombaméreg, az α-amanitin jelenlétében fejezik ki (1. táblázat). Érdekes módon az Amanita phalloides, a Death Cap gomba által termelt α-amanitin nagyon eltérően befolyásolja a három polimerázt. Az RNS polimeráz I teljesen érzéketlen az α-amanitinre, ami azt jelenti, hogy a polimeráz in vitro átírhatja a DNS-t a méreg jelenlétében. Ezzel szemben az RNS polimeráz II rendkívül érzékeny az α-amanitinre, az RNS polimeráz III pedig mérsékelten érzékeny., A transzkripciós polimeráz ismerete segíthet a kutatónak a vizsgált gén általános funkciójában. Mivel az RNS polimeráz II a gének túlnyomó többségét átírja, az eukarióta transzkripciós faktorokról és promoterekről folytatott későbbi megbeszéléseink során erre a polimerázra fogunk összpontosítani.
RNS polimeráz II promoterek és transzkripciós faktorok
Az eukarióta promoterek sokkal nagyobbak és bonyolultabbak, mint a prokarióta promoterek. Mindkettő azonban hasonló a prokarióták -10 szekvenciájához., Az eukariótákban ezt a szekvenciát TATA doboznak nevezik,és a kódolási szálon a konszenzus szekvencia TATAAA. Az eljárás megindításának (+1) helyéhez képest -25-35 bázison helyezkedik el (2.ábra). Ez a szekvencia nem azonos az E. coli -10 dobozzal, de megőrzi az A–T gazdag elemet. Az A–T kötések termosztálhatósága alacsony, ami segít a DNS-sablonnak lokálisan lazulni az átírásra való felkészülés során.,
Ahelyett, hogy az egyszerű σ tényező, amely segít a bind a prokarióta RNS polimeráz, hogy a szervező, eukaryotes össze egy komplex transzkripciós faktor szükséges toborozni RNS polimeráz II., hogy a fehérjét kódoló gént. A promoterhez kötődő transzkripciós tényezőket bazális transzkripciós tényezőknek nevezzük. Ezeket az alaptényezőket tfii-nek (transzkripciós faktor/polimeráz II), valamint egy további betűnek (a-J) nevezik. A magkomplexum a TFIID, amely TATA-kötő fehérjét (TBP) tartalmaz., A többi transzkripciós tényező szisztematikusan a DNS-sablonon helyezkedik el, mindegyik tovább stabilizálja az eljárás megindítását megelőző komplexet, és hozzájárul az RNS polimeráz II.
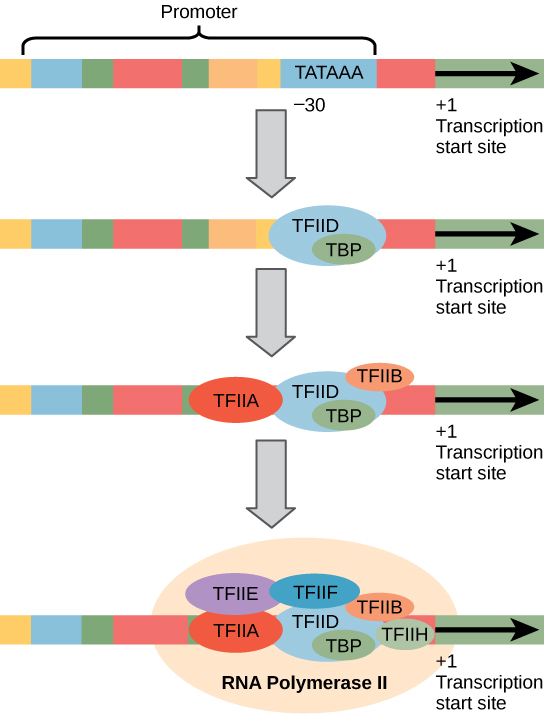
2.ábra. Az RNS polimeráz II által átírt gén generalizált promoterje látható. A transzkripciós tényezők felismerik a promoter-t. Az RNS polimeráz II ezután megköti és alkotja a transzkripciós iniciációs komplexet.,
practice Question
egy tudós egy eukarióta promoter-t egy bakteriális gén elé helyez, és a gént egy bakteriális kromoszómába helyezi. Elvárná, hogy a baktériumok átírják a gént?
néhány eukarióta promoternek is van egy konzervált CAAT box (GGCCAATCT)körülbelül -80. A Tata box előtt az eukarióta promóterek egy vagy több GC-ben gazdag dobozt (GGCG) vagy octamer dobozokat (ATTTGCAT) is tartalmazhatnak., Ezeket az elemeket köti mobil tényezők, amelyek növelik a hatékonyságot átírás kezdeményezése, gyakran azonosították a még “aktív” gének, amelyek folyamatosan fejezi ki a cellából.
a bazális transzkripciós faktorok kulcsfontosságúak egy PREINITÁCIÓS komplex kialakulásában a DNS-sablonon, amely később RNS polimeráz II-t toboroz a transzkripciós beavatáshoz. Az eukarióta transzkripció összetettsége nem ér véget a polimerázokkal és promoterekkel., Egy sereg más transzkripciós faktorok, amelyek kötődnek upstream erősítők és hangtompítók, is segít szabályozni a frekvencia, amellyel pre-mRNS szintetizálódik egy gén. A hangtompítók és hangtompítók befolyásolják a transzkripció hatékonyságát, de nem szükségesek a transzkripció folytatásához.
a promoterek fejlődése
a gének fejlődése ismerős fogalom lehet. A DNS-replikáció során génekben mutációk fordulhatnak elő, az eredmény a sejt számára előnyös lehet vagy nem., Egy enzim, szerkezeti fehérje vagy más tényező megváltoztatásával a mutáció folyamata átalakíthatja a funkciókat vagy a fizikai jellemzőket. Ugyanakkor az eukarióta promoterek és más génszabályozási szekvenciák is fejlődhetnek. Vegyünk például egy gént, amely sok generáció alatt értékesebbé válik a sejt számára. Talán a gén olyan szerkezeti fehérjét kódol, amelyet a sejtnek egy bizonyos funkcióhoz bőségesen kell szintetizálnia. Ha ez a helyzet, előnyös lenne a sejt számára, hogy a gén promoter toborozni transzkripciós faktorok hatékonyabban és növeli a gén expresszióját.,
a promoter-szekvenciák alakulását vizsgáló tudósok eltérő eredményekről számoltak be. Részben azért, mert nehéz pontosan megállapítani, hogy hol kezdődik és ér véget egy eukarióta promoter. Egyes promoterek géneken belül fordulnak elő; mások nagyon messze vannak a szabályozó gének upstream vagy akár downstream szintjén. Amikor azonban a kutatók korlátozott a vizsgálat, hogy az emberi alapvető szervező sorozatok meghatározott kísérletileg, mint a sorozatok, amik összekötik a preinitiation komplex, azt találták, hogy a támogatók fejlődni még gyorsabb, mint a fehérje kódoló gének.,
még mindig nem tisztázott, hogy a promoter evolúció hogyan felel meg az emberek vagy más magasabb szervezetek evolúciójának. Azonban egy promoter evolúciója, hogy egy adott génterméket többé-kevésbé hatékonyan készítsen, érdekes alternatíva a gének evolúciójához.
Szervező Struktúrák RNS Polimeráz i., III.
A folyamatok, hogy RNS polimeráz i., III., hogy a DNS-sablon jár valamivel kevesebb, komplex gyűjteménye transzkripciós faktor, de az általános téma ugyanaz.,
A kézirattár szervező elemei gének írta polimeráz i., III eltérnek által átírt RNS polimeráz II. RNS polimeráz én transcribes gének két GC-gazdag szervező szekvenciák a -45 +20 régióban. Ezek a szekvenciák önmagában elegendő átírás beavatás fordul elő, de a szervezők további szekvenciák a régióban a -180, hogy megfelelő -105 elé a beavatás oldalon tovább erősíti a beavatás. Az RNS-polimeráz III által transzkripált gének upstream promoterekkel vagy promoterekkel rendelkeznek, amelyek maguk a génekben fordulnak elő.,
Az eukarióta transzkripció egy szorosan szabályozott folyamat, amely különféle fehérjéket igényel, hogy kölcsönhatásba lépjenek egymással és a DNS-szálral. Bár az eukariótákban a transzkripció folyamata nagyobb metabolikus befektetést jelent, mint a prokariótákban, biztosítja, hogy a sejt pontosan átírja a fehérjeszintézishez szükséges pre-mRNS-eket.,
Nyúlás Megszűnése
a Következő megalakult a preinitiation komplex, a polimeráz megjelent a más transzkripciós faktorok, valamint nyúlás engedélyezett kell eljárni, mint a prokaryotes a polimeráz szintetizáló pre-mrns 5′ 3′ irányba. Amint azt korábban említettük, az RNS polimeráz II átírja az eukarióta gének nagy részét, így ez a szakasz arra összpontosít, hogy ez a polimeráz hogyan valósítja meg a nyúlást és a megszüntetést.,
bár a nyúlás enzimatikus folyamata lényegében azonos az eukariótákban és a prokariótákban, a DNS-sablon összetettebb. Amikor az eukarióta sejtek nem osztódnak, génjeik a DNS és a fehérjék diffúz tömegében, úgynevezett kromatinban léteznek. A DNS szorosan csomagolt körül töltött hiszton fehérjék ismételt időközönként. Ezeket a DNS–hiszton komplexeket, amelyeket együttesen nukleoszómáknak neveznek, rendszeresen elosztják, és 146 DNS-nukleotidot tartalmaznak, amelyek nyolc hiszton körül vannak, mint egy tekercs körül.,
ahhoz, hogy a polinukleotid szintézis megtörténjen, a transzkripciós gépnek el kell távolítania a hisztonokat az útból minden alkalommal, amikor nukleoszómával találkozik. Ezt egy FACT nevű speciális fehérje komplex végzi ,amely ” megkönnyíti a kromatin transzkripciót.”Ez a komplex elhúzza a hisztonokat a DNS-sablontól, amikor a polimeráz mozog rajta. Miután a pre-mRNS szintetizálódik, a TÉNYKOMPLEX helyettesíti a hisztonokat, hogy újra létrehozzák a nukleoszómákat.
a transzkripció megszűnése a különböző polimerázok esetében eltérő., A prokariótákkal ellentétben az eukariótákban az RNS-polimeráz II megnyúlása 1000-2000 nukleotiddal történik az átírandó gén végén túl. Ezt a pre-mRNS farkát az mRNS feldolgozás során hasítással távolítják el. Másrészt az I. és III. RNS polimerázok végződési jeleket igényelnek. Az RNS polimeráz I által átírt gének egy specifikus 18-nukleotid szekvenciát tartalmaznak, amelyet egy végprotein ismer fel. Az RNS polimeráz III végződésének folyamata egy mRNS hajtűt foglal magában, amely hasonló a prokarióták transzkripciójának Rho-független megszüntetéséhez.,
próbálja ki
hozzájárul!
javítsa ezt az oldalatismerjen meg többet
- H Liang et al., “Fast evolution of core promoters in primate genomes,” Molecular Biology and Evolution 25 (2008): 1239-44. ↵















