Határok a Növény Tudományos
Bevezető
Helyes progresszió a gyümölcs érési eléréséhez elengedhetetlen, hogy mindkét optimális gyümölcs minőség, hosszú élettartam, fontos tulajdonságok, amelyek meghatározzák ár piacok, illetve a végső nyereség. Az érés azonban összetett folyamat, amelyben számos tényező érintett, beleértve a hormonális kontrollt is, amely szabályozza a biokémiai és fiziológiai változásokat, amelyek végső érzékszervi és táplálkozási gyümölcs tulajdonságokat adnak., Hagyományosan a húsos gyümölcsöket az érlelési folyamat szerint klimatikus és nem klimatikus gyümölcsökké minősítették. A klimatikus gyümölcsök, mint például a paradicsom vagy a banán, a légzési sebesség és az etilén-termelés növekedését igénylik, hogy autokatalitikus válasz esetén felszabadítsák az érési folyamatot (Barry and Giovannoni, 2007; Liu et al., 2015)., Antagonistically, nem klimax gyümölcsök, mint a citrusfélék vagy a szőlőt nem mutat egy sorozatot sem etilén, sem a légzés előtt érés kezdete, pedig ezek a gyümölcsök, érési elsősorban által ellenőrzött progresszív felhalmozódása a phytohormone abszcizinsav esetében (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Mindazonáltal az új tanulmányok megerősítik azt az elképzelést, hogy az érési folyamatot nemcsak egy fitohormon termelése szabályozza, hanem inkább úgy tűnik, hogy egy ellenőrzött hormonális egyensúly eredménye (Symons et al., 2012; Teribia et al., 2016; Li et al., 2019).,
az Auxinok, a szalicilsav (SA) és a melatonin olyan fitohormonok, amelyek részt vesznek a növények számos kulcsfontosságú folyamatának jelzésében és szabályozásában. Az auxinokat széles körben növekedési és fejlesztési szabályozókként írták le, több funkcióval a növényekben (lásd Taylor-Teeples et al., 2016); az SA kiváltja a biotróf és hemi-biotróf kórokozók elleni védekezési választ (Loake and Grant, 2007), valamint fontos szerepet játszik az abiotikus stressz (Dong et al., 2014; Wani et al., 2017), virágzási és sejtciklus-kontroll (Carswell et al., 1989; Eberhard et al.,, 1989); és a melatonin nem csak azt találták, hogy auxin-szerű funkciók (Chen et al., 2009; Zuo et al., 2014; Wen et al., 2016), de azt is javasolták, hogy bizonyos növényekben potenciális antioxidánsként működjön (Arnao and Hernández-Ruiz, 2015), valamint a kórokozókra adott növényi válaszok szabályozójaként (Chen et al., 2018). Érdekes, hogy ez a három hormon közös prekurzor-chorismate -, így metabolikus kereszt-beszélgetés történik közöttük, és számos gént kell finoman szabályozni, hogy elterelje chorismate anyagcsere felé ezeket a vegyületeket., Az elmúlt években számos vélemény foglalkozott az auxin bioszintézisével és szerepével (Paul et al., 2012), SA (Asghari és Aghdam, 2010) és melatonin (Feng et al., 2014; Arnao and Hernández-Ruiz, 2018) in fruits. Bioszintézisüket és funkcióikat azonban többnyire külön írták le. Itt az auxin, SA és melatonin bioszintézisének, anyagcseréjének és funkciójának közös és differenciált aspektusait fogjuk megvitatni a klimatikus és nem klimatikus gyümölcsök növekedésében és érésében., Egy hangsúly lesz metabolikus elterelés főbb pontok a bioszintézis a chorismate, a szabályozó szerepet a érése klimax, mind a nem-klimax gyümölcsök, szerepük a moduláció a bioszintézis a bioaktív vegyületek, amely nagymértékben meghatározza a gyümölcs minőségét.
Chorismate-eredetű fitohormonok
a Chorismate a shikimate út végterméke, és kulcsszerepet játszik a fitohormonok bioszintézisében (1.ábra)., A korismate aromás aminosavakat, köztük triptofánt eredményez, és számos reakció révén, beleértve a triptofán indol-3-piruvinsavvá történő átalakítását a triptofán-aminotranszferáz (TA) által; az auxin indol-3-ecetsavat (IAA) képes előállítani. TA javasolták, mint egy univerzális kulcsfontosságú enzim IAA bioszintézis nem csak a vegetatív szervek (Enríquez-Valencia et al., 2018), de a reproduktív szervek fejlődésében is (Reyes-Olalde et al., 2017), beleértve a gyümölcsnövekedést és érést (Estrada-Johnson et al., 2017)., Ezzel szemben, amikor az izokoriszmát-szintáz (ICS) aktiválódik, a koriszmát izokoriszmáttá alakítható, az utóbbi pedig SA-ra alakítható. Annak ellenére, hogy ezt az utat először baktériumokban azonosították, jelenleg is jól megalapozott a növényekben (Wildermuth et al., 2001; Uppalapati et al., 2007; Catinot et al., 2008; Garcion et al., 2008; Abreu és Munné-Bosch, 2009). Végül a triptofán triptaminná alakulhat a triptofán dekarboxiláz (TDC) enzim segítségével., Bár ez az IAA bioszintézisének egy másik módja (a triptamin indol-3-acetaldehidré, majd később IAA-ba fordul), a TDC-t javasolták, hogy legyen az első sebességkorlátozó enzim a melatonin útvonalon (Kang et al., 2007). A TDC-t először az Apocynaceae családban azonosították (de Luca et al., 1989) később számos növényrendszerben (Byeon et al., 2012, 2014; Zhao et al., 2013; Wei et al., 2018)., A dekarboxilezés után a triptamint a triptamin-5-hidroxiláz szerotoninná alakítja, majd a szerotonin N-acetil-szerotoninná alakul át, hogy végül melatonint adjon a citoszolban (1.ábra).
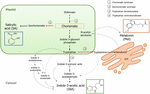
1. ábra. Az indol-3-ecetsav (IAA), a szalicilsav (SA) és a melatonin (Mel) bioszintézise a korismátból. Mindhárom vegyületnek közös végső prekurzora van (chorismate, narancssárga négyzetben) a shikimate útvonalból., A nagyobb eltérítési pontokat piros nyilakkal jelölik, a megfelelő enzimeket pedig a shikimáttól való eltérési sorrendben számozzák. A merész szaggatott nyilak egynél több lépést jeleznek a vegyületek között. A vékony szaggatott nyilak olyan útvonalakat jeleznek, amelyek még mindig nem jól definiáltak.
a chorismate-útvonal hormonok szintéziséhez vezető kulcs-átirányító enzimeket nemrégiben azonosították húsos gyümölcsökben (1.ábra). ICS, amely kifejezés kimutatták, hogy döntő fontosságú a biotikus stressz tolerancia (Garcion et al.,, 2008; Zeng és He, 2010), azonosították a paradicsomban és az almában. Zhu et al. (2016) jelentette az ICS1 (isochorismate synthase 1) túlexpresszióját hideg tárolási körülmények között paradicsomban, míg Zhang et al. (2017) talált aktiválása transzkripciós faktor részt vesz a kórokozóval kapcsolatos SA jelátviteli útvonal indukáló ICS kifejezés alma. Fokozott TDC-aktivitásról számoltak be éretlen paprika gyümölcsben kórokozók által okozott fertőzés esetén-fokozott TDC1 és TDC2 expresszió (Park et al., 2013) – az eperfa gyümölcsének növekedési szakaszában (Wang et al., 2016)., Végül a TA – t a grapevine-ban azonosították, mind a pre -, mind a véraison (Böttcher et al., 2013; Gouthu és Deluc, 2015, illetve az eper érése során (Estrada-Johnson et al., 2017). Figyelemre méltó azonban, hogy a gének, valamint enzimek leírt klimax gyümölcsök (azaz ICS) nem azonosítottak a nem-klimax gyümölcsök (azaz a TDC s TA), illetve fordítva; ezért további vizsgálatok elengedhetetlenek, hogy töltse ki ezeket a hiányzó ismeretek, illetve jobban megérteni, hogy ezek az elterelés pontokat közösen szabályozott során a gyümölcs érési.,
A Korismate-eredetű fitohormonok szerepe a klimatikus és nem klimatikus gyümölcsökben
a gyümölcsfejlődés mechanizmusainak feltárása a legutóbbi agronómiai kutatások egyik legfontosabb kihívása volt gazdasági következményei szempontjából. Ebben az összefüggésben phytohormones felhívták a figyelmet, mint felelős vezetők, a gyümölcs érése, kifejezetten etilén, illetve ABA a klimax, mind a nem-klimax gyümölcsök, ill. Azonban, hogy ezek a fitohormonok önmagukban szabályozhatják a gyümölcsfejlesztést, hamarosan túl egyszerűnek bizonyult., A kiterjedt kutatások, valamint az analitikai kémia és molekuláris technikák javulása után számos más hormon is megerősítést nyert, mint a gyümölcsfejlődés és érés potenciális szabályozói, beleértve a korismate-eredetű fitohormonokat is.
Auxins Cross-Talk Más Hormonok Során Gyümölcs Készlet, Növekedés, Érés
Auxins egy csoportja, a növényi hormonok, hogy alapvető szerepet játszanak a gyümölcs fejlődése, mind a nyomást gyakorolni a saját befolyásukat moduláló kifejezése más phytohormones., Az IAA endogén tartalma különösen magas a gyümölcskészletekben és a kezdeti növekedési fejlődési szakaszokban, majd az IAA mennyisége az érés kezdete előtt csökken, mind klimatikus (Zaharah et al., 2012) és a nem klimaxos gyümölcsök (Symons et al., 2012; Teribia et al., 2016), látszólag néhány kivétellel, mint az őszibarack (Tatsuki et al., 2013) és néhány szilvafajta (El-Sharkawy et al., 2014; ábra 2a). Bebizonyosodott, hogy az IAA részt vesz a gyümölcshalmazok gibberellinekkel (Mezzetti et al., 2004; Serrani et al., 2010; Bermejo et al., 2018; Hu et al.,, 2018). Az IAA bioszintézisének vagy jelátvitelének károsodása általában gyümölcs parthenokarpiához vezet, bár egyes gyümölcsökben rendellenes érést is eredményezhet (Wang et al., 2005; Liu J. et al., 2018; Reig et al., 2018). Magas tartalma iaa-n a kezdeti szakaszában a gyümölcs fejlődés előmozdítása gyümölcs növekedési miatt azonosítására vonzatok a sejtosztódás együttesen citokininek, valamint a kontroll sejt terjeszkedés kombinálva gibberellins (Liao et al., 2018)., Ebben az időszakban az auxinok és a gibberellinek közötti hormonális áthallás emellett lehetővé teszi a normál gyümölcsalakítást egy finomhangolt szabályozásban, amelyet az Auxin Válaszfaktorok közvetítenek (ARFs; Liao et al., 2018; Liu S. et al., 2018).
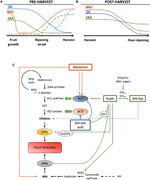
2.ábra. Az IAA, SA és Mel szerepe a klimatikus és nem klimatikus gyümölcsök fejlődése során. Modell, amely összefoglalja az IAA, SA és Mel kölcsönhatásait a klimatikus és nem klimatikus gyümölcsök érése során az A) elő-és B) a betakarítás után., Szaggatott vonalak jelzik a fitohormon tartalmának alternatív dinamikáját egyes gyümölcsökben (lásd a vita szövegét). C) az IAA, SA és Mel kölcsönhatásának áttekintése etilénnel és abszciszsavval (Aba) a klimatikus és nem klimatikus gyümölcsökben történő bioszintézissel. Az auxin az etilén bioszintézis pozitív szabályozója az ACC-szintáz gének (ACS) aktiválásával és az etilénre reagáló faktorok (ERFs) expresszióját indukálva. Az auxin 9-cis-epoxikarotenoid dioxigenáz (NCED) elnyomásával és az ABA-reagáló stresszgének (ARS) arf2 protein általi gátlásával is gátolja az ABA-termelést., Az endogén IAA-tartalom csökkenthető GH3 fehérjékkel, amelyek szintézisét etilén, ABA és cukrok segítik elő. A nem folytonos vonal jelzi az IAA IAA-Asp lehetséges szerepét, mint az etilént fokozó érési tényezőt egyes gyümölcsökben. A Melatonin fokozza az ABA és az etilén termelését, bár az ACS a melatonin tartalmától függően gátolható. Az SA az ACC-oxidáz (ACO) gének etilén elnyomásának gátlójaként működik, bár elősegítheti az ACS expresszióját., MTA, 50-metiltioadenozin; S-AdoMet, S-adenozil-metionin; SAM szintáz, S-adenozil-metionin szintáz; ACC, 1-aminociklopropán-1-karbonsav; IPP, izopentenil-difoszfát.
az endogén IAA-tartalom csökkenése az érés kezdete előtt több gyümölcsben is előfordult. Ezek a csökkentések az IAA-amido szintázok, a GH3-1 és a GH3-2 aszparaginsavval (IAA-Asp) való konjugációjával (2C.ábra) kapcsolatosak., Sőt, a GH3-1-GH3-2 mutatott magasabb kifejezés korai gyümölcs fejlődés, különösen az érés során megkezdését, mind a klimax gyümölcsök, mint például a paradicsom (Sravankumar et al., 2018) és az alma (Onik et al., 2018), valamint a nem klimatikus gyümölcsökben, mint a szőlőbogyók (Böttcher et al., 2010, 2011) és málna (Bernales et al., 2019)., Érdekes módon a szőlőbogyók fokozott GH3-1-expressziót mutattak az ABA és az etefon alkalmazása után, ami megmagyarázhatja az etilén bevonását az IAA tartalmának ellenőrzésébe az érés kezdete után, még a nem klimatikus gyümölcsökben is (Böttcher et al., 2010). Valójában számos tanulmány kiemeli az auxinok és az etilén szoros kölcsönhatását a gyümölcsérés során, kölcsönös hatással közöttük (Tadiello et al., 2016A; Busatto et al., 2017)., A klimatikus gyümölcsök esetében az IAA megnövekedett tartalma szükséges az ACC-szintáz gének (ACS) expressziójának aktiválásához, az ACC-szintetáz kódolásához, ami viszont az érési folyamatot kiváltó etilén termeléshez vezet (2C ábra; Tatsuki et al., 2013). Gyümölcsök, mint a paradicsom (Li et al., 2016) vagy banán (Choudhury et al.,, 2008), amelyek általában az érés kezdete előtt csökkent IAA-tartalmat tapasztalnak, érési késleltetést mutatnak, amikor az IAA-t vagy analógokat exogén módon alkalmazzák, míg az olyan gyümölcsök, mint az őszibarack, amelynek IAA-tartalma fokozatosan növekszik, amíg teljesen meg nem érik, gyorsított érést mutatnak az auxinok alkalmazásakor (tadiello et al., 2016b). Ezért az auxin-etilén kölcsönhatások szoros, összetett és differenciál szabályozásának léteznie kell a különböző gyümölcsökben a preharvest érés során., Ezenkívül az IAA-tartalom néhány nem klimatikus gyümölcs érése előtt csökken a 9-cis-epoxycarotenoid dioxigenáz (NCED) elnyomására és az ABA szintézis megkezdésére (2C.ábra; Jia et al., 2016). Érdekes módon ezt a folyamatot az IAA konjugáció is közvetíti fokozott GH3-1 aktivitással (Böttcher et al., 2010). Mindazonáltal Estrada-Johnson et al. (2017) kimutatta, hogy bár az iaa-n tartalmának csökkenése során eper érik, kifejezése FaAux/iaa-n, valamint FaARF gén család okozta a vörös tartályok, ami arra utal, bevonása azonosítására jelzés a teljesen érett gyümölcsök.,
a betakarítás után az auxin-tartalom általában változatlan marad, vagy általában csökken az oxidatív folyamatok miatt, amelyek az IAA endogén tartalmának kicsi, de progresszív csökkenését eredményezhetik (2b ábra). Az auxin kezelések a betakarítás után késleltetik a túlérést néhány gyümölcsben (Chen et al., 2016; Moro et al., 2017) és egyes szerves savak tartalmának növelése, a gyümölcs savasságának fenntartása (Li et al., 2017), ami arra utal, hogy az auxin szintén jelentős szerepet játszik a betakarítás utáni gyümölcsérés ellenőrzésében.,
Több Szerepet szalicilsav Során Gyümölcs Fejlődés, Érés
a szalicilsav egy másik chorismate eredetű phytohormone, hogy már többnyire kapcsolódik a védő hatása alatt biotikus stressz kontroll preharvest, illetve betakarítás utáni veszteséget származó kórokozó gyümölcs fertőzés (Babalar et al., 2007; Cao et al., 2013). Általában a szabad SA endogén tartalma magasabb a gyümölcsfejlődés kezdetén, majd fokozatosan csökken (Oikawa et al., 2015). Lu et al. (2015) az őszibarack gyümölcsének második növekedési szakaszában az SA másodlagos növekedését jelentette., Az SA banánban történő exogén alkalmazása késleltette az érési folyamatot, csökkentve a légzésrobbanást és a cellulóz-héj arányt, valamint a sejtfal lebomlásához és az antioxidáns rendszerhez kapcsolódó enzimek (Srivastava és Dwivedi, 2000) csökkenő aktivitását. Ezenkívül a cseresznyefák szalicilsavval és acetilszalicilsavval (ASA) történő kezelése kereskedelmi stádiumban megnövelte a cseresznye súlyát, szilárdságát és színét (Giménez et al., 2014). A sa vagy metil-szalicilát (MeSA) exogén alkalmazása a betakarítás utáni időszakban, késleltetett túlérés olyan gyümölcsökben, mint a kiwis (Zhang et al.,, 2003) és az édes cseresznye (Valero et al., 2011). Továbbá, kezelések szalicilsav vagy ASA enyhítette hűtés sérülés gránátalma (Sayyari et al., 2011), paradicsom (Aghdam et al., 2014) és avokádó (Glowacz et al., 2017). Az SA az ACC-oxidáz (ACO) expresszió gátlásával is kölcsönhatásba lép az etilénnel (Shi and Zhang, 2012). Meglepő módon az ACS expressziójának megnövekedett tartalma a körtében is megtalálható az SA alkalmazása után dózisszabályozott módon (Shi et al., 2013; Shi and Zhang, 2014). Ezért az etilén termelés SA-tól függő szabályozása finoman szabályozott (2C ábra).,
Melatonin: a Gyümölcsfejlődés szabályozásának feltörekvő szabályozója
a Melatonin a chorismate-eredetű út növényi hormonja is, amely az utóbbi években nagy figyelmet kapott (Hernández-Ruiz et al., 2005; Arnao and Hernández-Ruiz, 2018; Sharif et al., 2018). A közelmúltban a melatonint a gyümölcskészlet fontos tényezőjeként mutatták ki, hasonlóan az IAA-hatásokhoz (2a ábra; Liu S. et al., 2018). Emellett megfigyelték ennek a fitohormonnak a cirkadián termelését (Kolář et al., 1997)., A fa érése során a melatoninnal kezelt szőlő nagyobb szinkronitást és nagyobb súlyt mutatott, amikor teljesen megérett (Meng et al., 2015). Sőt, egy újabb tanulmány Xu et al. (2018) leírta, hogy a melatonin elősegítheti a szőlőbogyó érését az ABA-val és etilénnel való kölcsönhatása, valamint a hidrogén-peroxid révén. Kimutatták, hogy a melatonin hűtési toleranciát biztosít a gyümölcs hideg tárolása során, amely úgy tűnik, hogy leginkább az antioxidáns aktivitásához kapcsolódik (Cao et al., 2018; Aghdam et al., 2019)., A betakarítás utáni időszakban a paradicsom 50 µM-es melatonin (Sun et al., 2015), ellentétben a körtével, ahol a melatonin exogén alkalmazása 100 µM-en gátolta az etilén termelést (Zhai et al., 2018), vagy banángyümölcs, ahol a melatonin 50-200 µM-en gátolta az ACS és ACO expresszióját (2C. ábra; Hu et al., 2017). Bár a melatonin alkalmazásának különböző hatásai az adagolási hatáshoz kapcsolódhatnak, további kutatásokra van szükség ahhoz, hogy jobban megértsük a melatonin szerepét a gyümölcsfejlődés szabályozásában.,
A Korismate-eredetű fitohormonok Keresztbeszélése a Gyümölcsfejlődés szabályozásában
a korismate-eredetű fitohormonok közötti hormonális áthallás a gyümölcsfejlődés és érés során jelentkezett. Breitel et al. (2016) megállapította, hogy az auxinok kölcsönhatásba léphetnek az SA-val az ARF2-en keresztül (2C Ábra), és hogy az ARF2 paradicsomban történő túlexpressziója alacsonyabb SA-tartalmat és jelentős érési késleltetést eredményezett., Ez azt jelzi, hogy az auxinok korlátozhatják az SA termelését a gyümölcsfejlődés során, ami magyarázhatja a gyümölcsnövekedés (az auxin által közvetített) és a biotikus védekezés aktiválása (az SA által közvetített) közötti kereskedelmet. Továbbá, az SA exogén alkalmazása a papaya gyümölcsökön több IAA gén megváltozott expresszióját eredményezte, némelyikük down-szabályozott, míg mások up-szabályozott (Liu et al., 2017)., Ebben az esetben, az eredmények nem voltak meggyőzőek, hogy valójában mi történik módon a vélelmezett cross-talk között azonosítására, valamint SA során a gyümölcs érése, további kutatás szükséges ezen vagy más klimax gyümölcsök. Mindenesetre egyértelmű, hogy az IAA és az SA szoros kapcsolatban állnak egymással, nemcsak metabolikusan, hanem funkcionálisan is. Végül számos tanulmány számolt be a melatonin IAA-ra és SA bioszintézisre és/vagy jelzésre gyakorolt hatásáról (Arnao and Hernández-Ruiz, 2018), bár tudomásunk szerint ezen vizsgálatok egyikét sem klimatikus, sem nem klimatikus gyümölcsökben nem végezték el.,
Moduláció a Bioaktív Vegyületek Bioszintézis által Chorismate Eredetű Phytohormones
Bioaktív vegyületek (beleértve a fenolos vegyületek, isoprenoids, valamint antioxidáns vitaminok) nem csak széles körben vizsgálták, mint felelős konkrét érzékszervi tulajdonságai élelmiszerek, hanem a védő hatását az emberi sejtek ellen oxidatív folyamatok, a fejlesztés, a neurodegeneratív vagy a szív-érrendszeri betegségek, illetve bizonyos típusú rák (Liu et al., 1999; Mueller et al., 2010; Sturgeon and Ronnenberg, 2010; Björkman et al., 2011)., Fenolos vegyületek jelentősen hozzájárulnak közlésével különleges ízek, mint a tanninok – felelős a keserűség, vagy összehúzó ízét bizonyos gyümölcsöt, színek, mint például a antocianin pigmentek – felelős piros, kék, lila gyümölcs színek (Croteau et al., 2000). A fenolos vegyületek emberi fogyasztásra való relevanciája a kardiovaszkuláris és központi idegrendszeri egészséggel kapcsolatos oxidatív folyamatok elleni védőhatással, valamint a gyomor-bél traktus rákos megbetegedéseinek csökkent kockázatával (Björkman et al., 2011)., A karotinoidok, az izoprenoidokhoz tartozó tetraterpének szerepet játszanak a foto-oxidatív folyamatok elleni védelemben, valamint szerves pigmentekként; felelősek a gyümölcsök narancssárga-sárga színéért (Tapiero et al., 2004). Az étrendi karotinoidokról úgy gondolják, hogy az antioxidánsként betöltött szerepük miatt egészségügyi előnyökkel járnak a szembetegségek és bizonyos típusú rákok kockázatának csökkentésében (Johnson, 2002). A C-és E-vitaminok, beleértve az aszkorbinsavat és a tokoferolokat is, alapvető táplálkozási minőségi tényezők az emberi biológiai aktivitással rendelkező gyümölcsökben (Miret and Müller, 2017)., Figyelembe véve ezeknek a bioaktív vegyületeknek a gyümölcsökben az egészséget elősegítő vegyületekként betöltött szerepét, felhalmozódásuk modulációja kiemelkedő fontosságú mind a klimatikus, mind a nem klimatikus gyümölcsökben (Vasconsuelo and Boland, 2007). Ezeknek a bioaktív vegyületeknek a többsége magas szinten halmozódik fel a gyümölcs érése során, amikor a kevésbé ízletes zöld gyümölcsök táplálkozási szempontból gazdag, színes és ízletes gyümölcsökké alakulnak (Daood et al., 1996; Ranalli et al., 1998; Dumas et al., 2003; Singh et al., 2011)., Azonban az érlelés megkezdését követően az érés nem akadályozható meg, és általában túléréshez vezet, ami negatívan befolyásolja a gyümölcs minőségét (Kumar et al., 2014). Ezért a betakarítás utáni gyümölcskárosítás minimalizálása a bioaktív vegyületek magas tartalmának megszerzése mellett továbbra is az egyik legnagyobb kihívás a megoldáshoz.
az Auxint a karotinoid bioszintézis fontos szabályozójaként azonosították a klimatikus paradicsomos gyümölcsök érése során. A paradicsom érése a klorofill lebomlásával és a xantofillok karotinokra (β-karotin és likopin) való áttérésével jár (Fraser et al., 1994)., Iaa-n jelenik meg, hogy késedelem paradicsom érés által elnyomja etilén több upstream karotinoid átiratok beleértve Psy, Ziso, Pds, Critiso, valamint Chlases 1-3 meg, másrészt, elősegítve a felhalmozási β-Lyc1, valamint Crtr-β1 átiratát, ami a magasabb tartalmát xantofillek, valamint klorofill-a (Su et al., 2015). Ezen kívül néhány nem-klimax gyümölcsök, például cseresznye, szőlő, bogyós azonosítására is úgy tűnik, hogy módosítsam antocianin bioszintézis, ezáltal ellenőrzési gyümölcs érési folyamatokat. Teribia et al., (2016) negatív korrelációt jelentett az IAA és az antocianin tartalom között, jelezve, hogy az antocianin felhalmozódása az IAA tartalmának csökkenésekor kezdődik. Ezenkívül a szintetikus auxin-szerű vegyület, a benzotiazol-2-oxiecetsav (BTOA) alkalmazása késleltette az antocianin bioszintézis enzimjeit, például a kalcon szintetázt és az UDP-glükózt elősegítő gének up-szabályozását:flavonoid 3-o-glükoziltranszferáz a szőlőbogyókban (Davies et al., 1997).,
Pre -, illetve betakarítás utáni kezelések, szalicilátok, beleértve a SA, valamint a származékos ASA, valamint MeSA számoltak be, hogy szabályozza a bioaktív vegyületek felhalmozódása, ami javította az antioxidáns aktivitás, mind a klimax, mind a nem-klimax gyümölcsök. Számos gyümölcs, mint például a szilva, cseresznye, sárgabarack ismert, hogy egy rövid élet, a gyors minőségromlás a betakarítás után. Ezért folyamatosan keressük azokat a kezeléseket, amelyek javítják és fenntartják a gyümölcsminőséget, különösen a jótékony hatású bioaktív vegyületek tartalmát., A szilva SA-val, ASA-val és MeSA-val történő kezelése jelentős mennyiségű aszkorbinsavat, antocianint és fenolvegyületet eredményezett mind betakarításkor, mind hosszantartó hideg tárolás után (40 nap 4°C-on; Martínez-Esplá et al., 2017). Hasonló eredmények figyelhetők meg az SA, ASA és a MeSA édes cseresznyével végzett preharvest-kezeléseknél (Giménez et al., 2014, 2015, 2017). Továbbá, a betakarítás utáni kezelés SA, ASA, vagy MeSA fenntartotta a teljes fenolos tartalmát, valamint antocianin tartalmát hideg tárolás során gránátalma (sayyari et al., 2011), édes cseresznye (Valero et al.,, 2011), cornelian cherry (Dokhanieh et al., 2013), és barackos (Wang et al., 2015). Ezek az eredmények arra utalnak, hogy a szalicilátok részt vehetnek a fenilalanin ammónia-lyáz aktiválásában, amely a fenolos vegyületek bioszintezikus útjában részt vevő fő enzim (Martínez-Esplá et al., 2017).
a közelmúltban kimutatták, hogy a melatonin szabályozza a gyümölcs érését és modulálja a bioaktív vegyületeket. Az őszibarack gyümölcsökben a melatoninnal végzett betakarítás utáni kezelések antioxidáns rendszerek aktiválásával fokozták a hűtési toleranciát., Hűtés sérülés történik őszibarack gyümölcsök alacsony hőmérsékleten történő tárolás során (2-7, 6°C között) jellemző pépes barnulás, rendellenes érés és nagyobb érzékenység bomlás (Lurie and Crisosto, 2005). Cao et al. (2018) arról számolt be, hogy a melatoninnal kezelt őszibarack a tárolás 21.és 28. napján a PpGME, PpGPP és PpGLDH aszkorbinsav bioszintézis gének nagyobb átirata bőséget mutatott, mint a kontroll, ami az aszkorbinsav tartalmának növekedését eredményezte., Ezenkívül a melatonin fokozta a g6pdh, SKDH és PAL aktivitását, amelyek a fenolos vegyületek bioszintézisének alapvető enzimjei. A szerzők azt sugallják, hogy a melatonin kezelés bizonyos mértékig védi az őszibarackot a hűtés sérülésétől azáltal, hogy kifejezetten aktiválja a fenolos vegyületek bioszintézisét (Gao et al., 2018). Ezenkívül egy címkamentes differenciálproteomikai elemzés feltárta a melatonin hatását a gyümölcs érésének elősegítésére és az antocianin felhalmozódására a paradicsomgyümölcsök betakarítása után (Sun et al., 2016)., A szerzők arról számoltak be, hogy az exogén melatonin nyolc enzimet emelt az antocianin útvonalhoz kapcsolódóan, beleértve többek között a flavonol-3-hidroxilázt, a flavanon-3 béta-hidroxilázt, az antocianidin-szintetázt/leucoantocianidin-dioxigenázt és az antocianidin-3-o-glükóziltranszferázt. Továbbá a betakarítás utáni kezelés melatonin fokozott teljes fenolok, valamint antociánokat az eper gyümölcsök (Aghdam, valamint Fard, 2016-ban), valamint a késleltetett veszteség a teljes fenolok, flavonoidok, valamint antociánokat a licsi gyümölcsök (Zhang et al., 2018)., A melatonin-kezelés növelte a fenolok, antocianinok és flavonoidok tartalmát a szőlőbogyókban (Xu et al., 2017). Továbbá, a melatonin számoltak be, hogy fokozza a likopin felhalmozódása, valamint etilén termelés paradicsom, ami arra utal, hogy a melatonin növelheti a likopintartalom által hatással etilén bioszintézis, valamint jelzés (Sun et al., 2015)., Az etilén részvételét a bioaktív vegyületek melatonin által indukált szabályozásában a szőlőbogyókban is megfigyelték, ahol az etilén kettős blokkkezelése a melatonin polifenoltartalomra gyakorolt csökkent hatását mutatta (Xu et al., 2017).
következtetés
megállapítást nyert, hogy a koriszmátból származó fitohormonok, beleértve az auxint, SA-t és melatonint, nemcsak közös prekurzoruk, hanem alapvető szerepet játszanak a gyümölcsnövekedés és érés szabályozásában., A metabolikus és funkcionális kereszt-talk közöttük, valamint más fitohormonok fordul elő, egy spatiotemporális módon, hogy finoman szabályozza a fejlődését klimaxos, nem klimaxos gyümölcsök. A klimaxos és nem klimaxos gyümölcsök dinamikájának különbségei azt bizonyítják, hogy a chorismate-eredetű hormonok reakciója nem univerzális, hanem erősen fajspecifikus. Ezenkívül a korismate-eredetű fitohormonok modulálják a bioaktív vegyületek felhalmozódását is, ezáltal befolyásolják a gyümölcs minőségét., További kutatásokra van azonban szükség, hogy jobban megértsük, hogyan (nem) más hormonok, mint például az etilén vagy ABA, állítsd át a bioszintézis tanulás kulcs az anyagcsere-elterelés pontok, (ii) hatnak egymásra, a funkcionális, valamint molekuláris szinten, valamint (iii) a közösen moduláló bioaktív vegyületek bioszintézis, s következésképp befolyással nem csak a gyümölcs, növekedés, érés, hanem a minőség a klimax, mind a nem-klimax gyümölcsök.
szerzői hozzájárulások
SM-B a kéziratot MP-L. MP-l, PM, MM és SM-B segítségével dolgozta ki és tervezte meg., MP-L készített számok segítségével PM. Minden szerző hozzájárult az ötletek megvitatásához, felülvizsgálta és jóváhagyta a végső kéziratot.
finanszírozás
ezt a munkát a katalán kormány által az SM-B-nek adott ICREA Academia-díj finanszírozta.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
Böttcher, C., Boss, P. K., and Davies, C. (2011)., Acyl szubsztrát beállítások egy iaa-n-legkevésbé szintetáz figyelembe eltérések szőlő (Vitis vinifera L.) bogyó érése által okozott különböző auxinic vegyületek fontosságára utaló azonosítására ragozás a növény fejlődését. J. Exp. Bot. 62, 4267–4280. doi: 10.1093/jxb/err134 P>PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., and Sivakumar, D. (2017)., A hidegen tárolt “Hass” avokádó postharvest minőségének fenntartása a zsírsavtartalom és összetétel megváltoztatásával természetes illékony vegyületek – metil-jasmonát és metil-szalicilát-alkalmazásával. J. Sci. Kaja. Agric. 97, 5186–5193. doi: 10.1002 / jsfa.8400
P > PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Zeng, W., and He, S. Y. (2010). A flagellin receptor flagellin-SENSING2 kiemelkedő szerepe az Arabidopsis Pseudomonas syringae pv paradicsom DC3000-re adott sztomatális válasz közvetítésében. Növényi Fiziol. 153, 1188–1198. doi: 10.1104 / PP.110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















