Biology for Majors I (日本語)
学習成果
- RNAポリメラーゼの役割について説明します
このセクションでは、転写におけるRNAポリメラーゼの 転写の各段階におけるRNAポリメラーゼの役割を学ぶために読んでください。,
転写の開始
DNAテンプレートに単独で結合することができる原核生物ポリメラーゼとは異なり、真核生物は転写因子と呼ばれるいくつかの他のタンパク質を必要とし、最初にプロモーター領域に結合し、次に適切なポリメラーゼを募集するのを助ける。
三つの真核生物RNAポリメラーゼ
真核生物mRNA合成の特徴は、原核生物のものより著しく複雑である。 真核生物は、五つのサブユニットからなる単一のポリメラーゼの代わりに、それぞれ10個以上のサブユニットからなる三つのポリメラーゼを持っている。, それぞれの真核生物ポリメラーゼはまた、それをDNA鋳型に持ち込むために別個の転写因子のセットを必要とする。
RNAポリメラーゼIは、リボソームRNA(rRNA)が転写され、処理され、リボソームに組み立てられる特殊な核下部構造である核小体に位置している(表1)。 RRNA分子は、細胞の役割を持っているが、タンパク質に翻訳されていないため、構造Rnaと考えられています。 Rrnaはリボソームの構成要素であり、翻訳プロセスに不可欠である。 RNAポリメラーゼIは5S rRNA分子を除くすべてのrrnaを合成する。, “S”の指定は、遠心分離中に粒子が堆積する速度を特徴付ける非加法値である”Svedberg”単位に適用されます。
| テーブル1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., 真核生物のプレmrnaは、転写後、翻訳前に広範な処理を受けます(図1)。 明確にするために、真核生物における転写および翻訳に関するこのモジュールの議論では、翻訳される準備ができている成熟した処理された分子のみを表すために”mrna”という用語を使用することになる。 RNAポリメラーゼIIは、真核生物の遺伝子の圧倒的多数を転写するための責任があります。

図1. 真核生物のmRNAは、スプライシングされなければならないイントロンを含む。 5’帽子および3’多尾はまた加えられる。,
RNAポリメラーゼIIIも核内に位置する。 このポリメラーゼは、5SプレrRNA、トランスファープレRna(プレtrna)、および小さな核プレRnaを含む様々な構造Rnaを転写する。 Trnaは翻訳において重要な役割を果たし、mRNAテンプレートと成長するポリペプチド鎖との間のアダプター分子として機能します。 小さな核Rnaは、”スプライシング”プレmrnaおよび転写因子を調節するなど、様々な機能を有する。,
新しい遺伝子を特徴付ける科学者は、遺伝子が特定のキノコ毒α-アマニチンの存在下で発現されているかどうかをテストすることによって、どのポリメラーゼがそれを転写するかを決定することができる(表1)。 興味深いことに,死のキノコであるアマニタファロイデスによって産生されるα-アマニチンは,三つのポリメラーゼに非常に異なる影響を与える。 RNAポリメラーゼIはα-アマニチンに完全に鈍感であり、ポリメラーゼはこの毒の存在下でin vitroでDNAを転写することができることを意味する。 対照的に、RNAポリメラーゼIIはα-アマニチンに対して非常に感受性であり、RNAポリメラーゼIIIは適度に感受性である。, 転写ポリメラーゼを知ることは、研究者が研究されている遺伝子の一般的な機能を手掛かりにすることができます。 RNAポリメラーゼIIは大部分の遺伝子を転写するため、真核生物の転写因子とプロモーターについてのその後の議論では、このポリメラーゼに焦点を当てます。
RNAポリメラーゼIIプロモーターと転写因子
真核生物のプロモーターは、原核生物のプロモーターよりもはるかに大きく、より複雑です。 しかしながら、両方とも原核生物の-10配列と同様の配列を有する。, 真核生物では、この配列はTATAボックスと呼ばれ、コーディング鎖上にコンセンサス配列TATAAAを有する。 それは開始(+25)サイトに対して-35塩基に位置しています(図2)。 この配列は大腸菌–10ボックスと同一ではありませんが、A-Tリッチ要素を保存します。 A-T結合の熱安定性は低く、これはDNAテンプレートが転写の準備のために局所的に巻き戻すのを助ける。,
原核生物のRNAポリメラーゼをそのプロモーターに結合するのに役立つ単純なγ因子の代わりに、真核生物はRNAポリメラーゼIIをタンパク質コード遺伝子にリクルートするために必要な転写因子の複合体を組み立てる。 プロモーターに結合する転写因子は基底転写因子と呼ばれる。 これらの基底因子はすべてTFII(転写因子/ポリメラーゼII)に加えて追加の文字(A-J)と呼ばれます。 コア複合体はTFIIDであり、これはTATA結合タンパク質(TBP)を含む。, 他の転写因子はdnaテンプレート上に体系的に配置され、それぞれが開始前複合体をさらに安定化させ、RNAポリメラーゼIIの募集に寄与する。
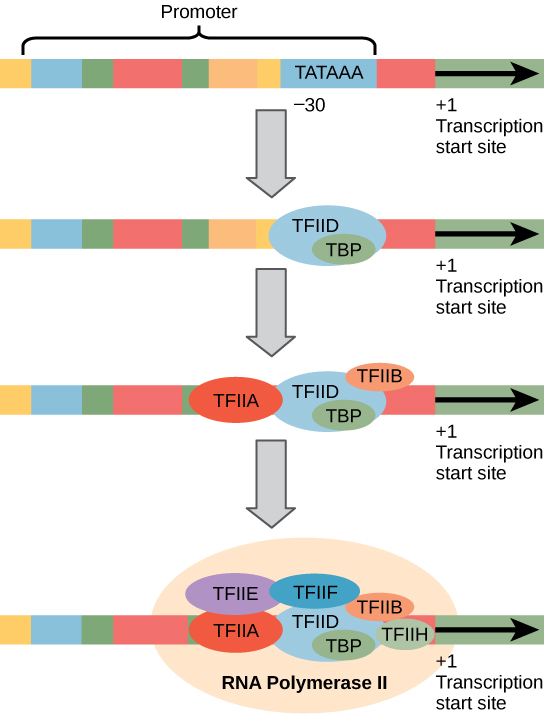
図2。 RNAポリメラーゼIIによって転写される遺伝子の一般化プロモーターを示した。 転写因子はプロモーターを認識する。 次にRNAポリメラーゼIIが結合して転写開始複合体を形成する。,
練習問題
科学者は、細菌遺伝子の前に真核生物のプロモーターをスプライスし、細菌染色体に遺伝子を挿入します。 あなたは細菌が遺伝子を転写すると期待しますか?
いくつかの真核生物のプロモーターはまた、約-80で保存されたCAATボックス(GGCCAATCT)を持っています。 TATAボックスのさらに上流では、真核生物のプロモーターはまた、一つ以上のGCリッチボックス(GGCG)または八量体ボックス(ATTTGCAT)を含有してもよい。, これらの要素は、転写開始の効率を高める細胞因子に結合し、しばしば細胞によって常に発現されているより”活性な”遺伝子において同定される。
基礎転写因子は、その後転写開始のためのRNAポリメラーゼIIを募集するDNAテンプレート上のpreinitiation複合体の形成に重要です。 真核生物の転写の複雑さは、ポリメラーゼおよびプロモーターで終わらない。, 上流のエンハンサーおよびサイレンサーに結合する他の転写因子の軍隊はまた、pre-mRNAが遺伝子から合成される頻度を調節するのを助ける。 エンハンサーおよびサイレンサーは転写の効率に影響を与えるが、転写が進行するためには必要ではない。
プロモーターの進化
遺伝子の進化はよく知られた概念であるかもしれません。 突然変異は、DNA複製中に遺伝子において起こり得、その結果は、細胞に有益であり得るか、または有益でないかもしれない。, 酵素、構造蛋白質、または他の要因の変更によって、突然変異のプロセスは機能か物理的な特徴を変形できます。 しかしながら、真核生物のプロモーターおよび他の遺伝子調節配列も同様に進化する可能性がある。 例えば、多くの世代にわたって、細胞にとってより貴重になる遺伝子を考えてみましょう。 たぶん遺伝子は、細胞が特定の機能のために豊富に合成する必要がある構造タンパク質をコードしているかもしれません。 この場合、その遺伝子のプロモーターが転写因子をより効率的に募集し、遺伝子発現を増加させることは、細胞にとって有益であろう。,
プロモーター配列の進化を調べる科学者は、さまざまな結果を報告しています。 部分的には、これは、真核生物のプロモーターがどこで始まり、終わるかを正確に推測することが困難であるためである。 いくつかのプロモーターは遺伝子内で起こり、他のプロモーターは、それらが調節している遺伝子の非常に上流、または下流に位置する。 しかし、研究者らは、開始前複合体に結合する配列として実験的に定義されたヒトコアプロモーター配列に検査を限定したとき、プロモーターはタンパク質コード遺伝子よりもさらに速く進化することを見出した。,
プロモーターの進化がヒトまたは他の高等生物の進化にどのように対応しているかはまだ不明です。 しかし、与えられた遺伝子産物の多かれ少なかれを効果的に作るためのプロモーターの進化は、遺伝子自体の進化に対する興味深い代替物である。
RNAポリメラーゼIおよびIIIのプロモーター構造
RNAポリメラーゼIおよびIIIをDNAテンプレートにもたらすプロセスは、転写因子のわずかに複雑なコレクションを含むが、一般的なテーマは同じである。,
ポリメラーゼIおよびIIIによって転写される遺伝子の保存されたプロモーター要素は、RNAポリメラーゼIIによって転写されるものとは異なります。RNAポリメラーゼIは、-45から+20領域にGCに富むプロモーター配列を持つ遺伝子を転写します。 これらの配列だけでは転写開始が起こるのに十分であるが、開始部位の上流の-180から-105までの領域に追加の配列を有するプロモーターは、開始をさらに高めるであろう。 RNAポリメラーゼIIIによって転写される遺伝子は、遺伝子自体の中で起こる上流のプロモーターまたはプロモーターを有する。,
真核生物の転写は、互いにおよびDNA鎖と相互作用するために様々なタンパク質を必要とする厳密に調節されたプロセスである。 真核生物における転写のプロセスは、原核生物よりも大きな代謝投資を伴うが、それは細胞がタンパク質合成に必要なプレmrnaを正確に転写するこ,
伸長および終了
前開始複合体の形成に続いて、ポリメラーゼは他の転写因子から放出され、ポリメラーゼが5’から3’方向にpre-mRNAを合成することにより、原核生物のように伸長が進行することができる。 前に説明したように、RNAポリメラーゼIIは真核生物の遺伝子の主要なシェアを転写するので、このセクションでは、このポリメラーゼがどのように伸長と終了を達成するかに焦点を当てます。,伸長の酵素的プロセスは、真核生物および原核生物において本質的に同じであるが、DNA鋳型は、より複雑である。 真核細胞が分裂していないとき、それらの遺伝子は、クロマチンと呼ばれるDNAおよびタンパク質の拡散塊として存在する。 DNAは繰り返された間隔で荷電したヒストン蛋白質のまわりで堅く包まれる。 これらのDNA–ヒストン複合体は、総称してヌクレオソームと呼ばれ、規則的に間隔をあけられ、スプールの周りの糸のような八つのヒストンの周りに巻かれた146,
ポリヌクレオチド合成が起こるためには、転写機構は、ヌクレオソームに遭遇するたびにヒストンを邪魔にならないように移動させる必要がある。 これは、FACTと呼ばれる特殊なタンパク質複合体によって達成され、”クロマチン転写を促進する。”この複合体は、ポリメラーゼがそれに沿って移動すると、DNA鋳型からヒストンを引き離します。 Pre-mRNAが合成されると、FACT複合体はヒストンを置き換えてヌクレオソームを再作成する。
転写の終了は、異なるポリメラーゼによって異なる。, 原核生物とは異なり、真核生物におけるRNAポリメラーゼIIによる伸長は、転写される遺伝子の終わりを超えて1,000–2,000ヌクレオチドを起こす。 このプレmrna尾部は、その後、mrna処理中の切断によって除去される。 一方、RNAポリメラーゼIおよびIIIは終了シグナルを必要とする。 RNAポリメラーゼIによって転写される遺伝子は、終端タンパク質によって認識される特定の18ヌクレオチド配列を含む。 RNAポリメラーゼIIIにおける終了のプロセスは、原核生物における転写のrho非依存的終了と同様のmRNAヘアピンを含む。,
試してみてください
貢献してください!
このページを改善してくださいもっと学ぶ
- H Liang et al.,”霊長類ゲノムにおけるコアプロモーターの高速進化,”Molecular Biology and Evolution25(2008):1239-44. li>















