생물학 전공자가
학습 결과
- 의 역할을 이해합니다 RNA
이 섹션에서 확장됩니다에 따라 특정한 역할의 RNA 폴 동안 전사. 전사의 각 단계에서 RNA 중합 효소의 역할을 배우기 위해 계속 읽으십시오.,
의 개시 전사
과는 달리 prokaryotic 효소 바인딩할 수 있는 DNA 템플릿에서 그것의 자신의,진핵생물에 필요한 여러 가지 다른 단백질이라는 전사 인 요소,처음에 바인딩하여 발기인 영역과한 다음 도움을 모집에 적합한 효소.
3 개의 진핵 생물 RNA 중합 효소
진핵 생물 mRNA 합성의 특징은 원핵 생물의 특징이 현저하게 더 복잡하다. 대신 하나의 중합효소 연 포함하는 다섯 subunits,은 진핵생물 세 폴는 각각의 10subunits 나 더 있습니다., 각각의 진핵 생물 중합 효소는 또한 DNA 템플릿으로 가져 오기 위해 뚜렷한 전사 인자 세트를 필요로한다.
RNA 중합 효소 I 는 리보솜 RNA(rRNA)가 전사되고,처리되고,리보솜으로 조립되는 특수 핵 하부 구조 인 nucleolus 에 위치한다(표 1). RRNA 분자는 세포 역할을 가지고 있지만 단백질로 번역되지 않기 때문에 구조적 Rna 로 간주됩니다. RRNAs 는 리보솜의 구성 요소이며 번역 과정에 필수적입니다. RNA 중합 효소 I 는 5S rRNA 분자를 제외한 모든 rrna 를 합성합니다., “S”지정은 원심 분리 중에 입자가 퇴적되는 속도를 특징 짓는 비 첨가 값인”Svedberg”단위에 적용됩니다.
| 표 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., 진핵 생물 pre-mRNAs 는 전사 후 그러나 번역 전에 광범위한 처리를 거친다(그림 1). 명확성을 위해,이 모듈의의 논의를 전사 및 번역 진핵생물에서 사용하는 용어”mRNAs 를 설명하는”성숙,분자 처리할 수 있는 번역하고 있습니다. Rna 중합 효소 II 는 압도적 인 대다수의 진핵 생물 유전자를 전사 할 책임이있다.

그림 1. 진핵 생물 mRNA 에는 접합해야하는 인트론이 들어 있습니다. 5’캡과 3’폴리-꼬리도 추가됩니다.,
RNA 중합 효소 III 는 또한 핵에 위치한다. 이 중합 효소는 5S pre-rRNA,transfer pre-RNAs(pre-tRNAs)및 작은 핵 pre-rnas 를 포함하는 다양한 구조적 RNAs 를 전사합니다. TRNAs 에는 번역에 있는 긴요한 역할이 있습니다;그들은 mRNA 템플렛과 성장하는 폴리펩티드 사슬 사이 접합기 분자로 봉사합니다. 작은 핵 Rna 는 pre-mRNAs 를”접합”하고 전사 인자를 조절하는 것을 포함하여 다양한 기능을 가지고있다.,
과학자 특성화 하는 새로운 유전자를 결정할 수 있는 효소 표여하는지 여부를 테스트하고 유전자 표현의 존재에서 특정한 버섯 독 α-amanitin(표 1). 흥미롭게도,데스 캡 버섯 인 amanita phalloides 에 의해 생성 된 α-amanitin 은 세 가지 중합 효소에 매우 다르게 영향을 미친다. RNA 중합 효소 I 는 α-amanitin 에 완전히 민감하지 않으므로 중합 효소가이 독의 존재 하에서 시험 관내에서 DNA 를 전사 할 수 있음을 의미합니다. 대조적으로,rna 중합 효소 II 는 α-amanitin 에 매우 민감하고,RNA 중합 효소 III 는 적당히 민감하다., 전사 중합 효소를 알면 연구자가 연구중인 유전자의 일반적인 기능에 단서를 줄 수 있습니다. 기 때문에 RNA polymerase II 표기 대부분의 유전자를 우리에 초점을 맞출 것이 이 효소에서 우리의 이후의에 대한 토론을 진핵세포 전사 요인과 발기인입니다.
RNA 중합 효소 II 프로모터 및 전사 인자
진핵 생물 프로모터는 원핵 생물 프로모터보다 훨씬 크고 복잡합니다. 그러나 둘 다 원핵 생물의 -10 서열과 유사한 서열을 가지고있다., 진핵생물에서 이 순서가라는 타타 상자,그리고 합의 순서 TATAAA 에 코딩드도 있습니다. 개시(+1)부위에 상대적인 -25 내지 -35 염기에 위치한다(그림 2). 이 서열은 대장균 -10 상자와 동일하지는 않지만 a–T 가 풍부한 원소를 보존합니다. A–T 결합의 thermostability 는 낮 이것은 DNA 템플렛이 전사를 위해 준비에서 국부적으로 푸는 것을 돕는다.,
대신에의 간단한 σ 요소 바인딩 할 수 prokaryotic RNA 의 발기인,진핵생물 조립의 복잡한 전사 하는 데 필요한 요소 모집 RNA polymerase II 단백질을 코딩하는 유전자. 프로모터에 결합하는 전사 인자를 기초 전사 인자라고합니다. 이러한 기저 인자는 모두 tfii(전사 인자/중합 효소 II 의 경우)와 추가 문자(A-J)라고합니다. 핵심 복합체는 TATA 결합 단백질(TBP)을 포함하는 TFIID 입니다., 다른 녹음 요인을 체계적으로 떨어지는 장소에서 DNA 템플릿으로,각각의 하나 더 안정시키 pre-initiation complex 고에 기여하고 채용의 RNA polymerase II.
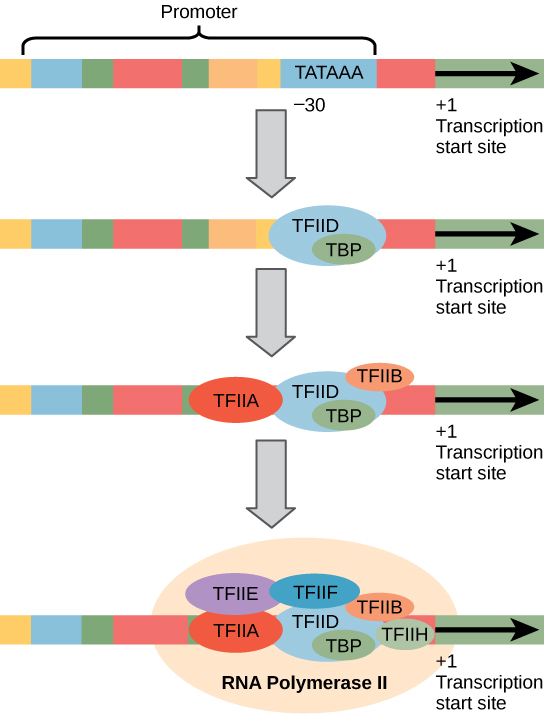
림 2. RNA 중합 효소 II 에 의해 전사 된 유전자의 일반화 된 프로모터가 도시되어있다. 전사 인자는 프로모터를 인식합니다. 그런 다음 rna 중합 효소 II 가 결합하여 전사 개시 복합체를 형성합니다.,
연습 문제
과학자가 결합 진 핵 프로모터에서 앞의 세균 유전자가 삽입된 유전자에 세균 염색체입니다. 박테리아가 유전자를 전사 할 것으로 기대하겠습니까?
일부 진핵 생물 프로모터는 또한 약 -80 에서 보존 된 CAAT 상자(GGCCAATCT)를 갖는다. TATA 박스의 더 상류에서,진핵 생물 프로모터는 또한 하나 이상의 GC-리치 박스(GGCG)또는 옥타머 박스(ATTTGCAT)를 포함 할 수있다., 이러한 요소 바인딩은 셀룰러 요소의 효율성을 증가 녹음방송 개시 및 종종에서 식별상”active”유전자는 지속적으로 표현합니다.
기초 전사 인자는 전사 개시를 위해 이후 rna 중합 효소 II 를 모집하는 DNA 템플릿에 preinitiation 복합체의 형성에 결정적이다. 진핵 생물 전사의 복잡성은 중합 효소 및 프로모터로 끝나지 않습니다., 상류 인핸서 및 소음기에 결합하는 다른 전사 인자의 군대는 또한 유전자로부터 pre-mRNA 가 합성되는 빈도를 조절하는 데 도움을줍니다. 인핸서 및 소음기는 전사의 효율에 영향을 미치지 만 전사가 진행되기 위해서는 필요하지 않습니다.
프로모터의 진화
유전자의 진화는 친숙한 개념 일 수있다. 돌연변이는 DNA 복제 과정에서 유전자에서 발생할 수 있으며 그 결과는 세포에 유익 할 수도 있고 그렇지 않을 수도 있습니다., 효소,구조 단백질 또는 다른 요인을 변경함으로써 돌연변이 과정은 기능이나 신체적 특징을 변형시킬 수 있습니다. 그러나,진핵 생물 프로모터 및 다른 유전자 조절 서열도 진화 할 수있다. 예를 들어,여러 세대에 걸쳐 세포에 더 가치있게되는 유전자를 생각해보십시오. 어쩌면 유전자는 세포가 특정 기능을 위해 풍부하게 합성해야하는 구조 단백질을 암호화합니다. 이 경우,도움이 될 것입니다에 대한 셀는 유전자 발기인을 모집하는 전사 인 요소를 더 효율적으로 증가하고 유전자 발현.,
프로모터 서열의 진화를 조사한 과학자들은 다양한 결과를보고했다. 부분적으로 이것은 진핵 생물 프로모터가 시작되고 끝나는 곳을 정확히 추론하기가 어렵 기 때문입니다. 일부 발기인에서 발생하는 유전자,다른 위치가 멀리 업스트림,심지어 다운스트림,유전자의 그들이 조절이다. 그러나 때,연구자들은 제한 그들의 심사를 인간의 핵심 프로모터 시퀀스가 정의된 실험적으로 시퀀스로는 바인딩 preinitiation 복잡,그들이 발견되는 발기인 진화보다 더 빨리 단백질을 코딩하는 유전자가 있습니다.,
프로모터 진화가 인간이나 다른 고등 생물의 진화에 어떻게 대응할 수 있는지는 여전히 불분명하다. 그러나,진화의 발기인을 효과적으로 만들은 더 많거나 적은 주어진 유전자의 제품은 흥미로운 대체하는 유전자의 진화 자체.
발기인 구조에 대한 RNA 폴 나와 III
프로세스의 데 RNA 폴 나와 III DNA 템플릿을 포함한 약간 덜 복잡한 컬렉션의 전사 인하지만,일반적인 주제는 동일합니다.,
보존된 발기인 요소를 위한 유전자 전사에 의해 폴 나와 III 다 transcribed by RNA polymerase II. RNA polymerase I 표 유전자가 있는 두 개의 GC-이 풍부한 프로모터 시퀀스에서 -45to+20 지역입니다. 이러한 시퀀스에 혼자 있는 충분한 전송 개시 발생하는지만,프로모터와 추가적인 시퀀스에서 이 지역에서 -180 을 -105 업스트림 개시의 사이트는 더욱 향상시됩니다. RNA 중합 효소 III 에 의해 전사되는 유전자는 유전자 자체 내에서 발생하는 상류 프로모터 또는 프로모터를 갖는다.,
진핵 생물 전사는 서로 및 DNA 가닥과 상호 작용하기 위해 다양한 단백질을 필요로하는 엄격하게 조절 된 과정입니다. 지만 전사 프로세스 진핵생물에서 포함 큰 대사보다는 투자에서 prokaryotes,그것은 세포 표을 정확하게 pre-mRNAs 하는데 필요한 단백질 합니다.,
신장 및 종료
은 다음의 형성 preinitiation 복잡한,효소가 발표에서 다른 녹음 요인,그리고 신장가를 진행할 수 있으로 prokaryotes 으로 효소를 합성하 pre-mRNA5’3’방향이다. 앞에서 설명한 대로,RNA polymerase II 표 주요 주의 진핵세포의 유전자,이 섹션에 초점을 맞출 것이 어떻게 이 효소를 수행한 신율 및 종료됩니다.,
신장의 효소 과정은 진핵 생물과 원핵 생물에서 본질적으로 동일하지만 DNA 템플릿은 더 복잡합니다. 진핵 세포가 분열하지 않을 때,그들의 유전자는 염색질이라고 불리는 DNA 와 단백질의 확산 덩어리로 존재합니다. DNA 는 반복 간격으로 하전 된 히스톤 단백질 주위에 단단히 포장됩니다. 이러한 DNA–히스톤 단지 라고 통칭 nucleosomes,은 간격의 정기적으로 포함 146 뉴클레오티드의 DNA 를 상처를 주는 여덟 히스톤 같은 스레드 스풀 주위에.,
뉴클레오티드에 대한 종합하여 발생,전송 기계에 이동해야 히스톤 방식의 모든 시간은 만남 nucleosome. 이것은 팩트라고 불리는 특별한 단백질 복합체에 의해 성취되며,이는”염색질 전사를 용이하게합니다.”이 복합체는 중합 효소가 그것을 따라 이동함에 따라 히스톤을 DNA 템플릿으로부터 멀리 끌어 낸다. Pre-mRNA 가 합성되면,FACT complex 는 히스톤을 대체하여 뉴 클레오 솜을 다시 만듭니다.
전사의 종결은 상이한 중합 효소에 대해 상이하다., 원핵 생물에서와는 달리,진핵 생물에서 RNA 중합 효소 II 에 의한 신장은 전사되는 유전자의 말단을 넘어 1,000–2,000 뉴클레오타이드가 일어난다. 이 pre-mRNA 꼬리는 mRNA 처리 동안 분열에 의해 연속적으로 제거된다. 반면에,RNA 중합 효소 I 및 III 는 종단 신호를 필요로한다. RNA 중합 효소 I 에 의해 전사 된 유전자는 종단 단백질에 의해 인식되는 특정 18-뉴클레오티드 서열을 함유한다. RNA 중합 효소 III 에서의 종결 과정은 원핵 생물에서의 전사의 rho-독립적 인 종결과 유사한 mRNA 머리핀을 포함한다.,나는 그것을 시도 할 수 없다.
이 페이지를 개선하십시오.
- H Liang et al.,”영장류 게놈에서 핵심 프로모터의 빠른 진화”,분자 생물학 및 진화 25(2008):1239-44. ↵















