Biologi för Majors i
läranderesultat
- beskriv rollen som RNA-polymeras
det här avsnittet kommer att expandera på rna-polymerasernas specifika roll under transkriptionen. Läs vidare för att lära sig rollen som RNA-polymeraser vid varje transkriptionsstadium.,
initiering av transkription
Till skillnad från prokaryotiskt polymeras som kan binda till en DNA-mall på egen hand kräver eukaryoter flera andra proteiner, kallade transkriptionsfaktorer, för att först binda till promotorregionen och sedan hjälpa till att rekrytera lämpligt polymeras.
de tre eukaryota rna-Polymeraserna
funktionerna i eukaryotisk mRNA-syntes är markant mer komplexa hos prokaryoter. I stället för ett enda polymeras bestående av fem underenheter har eukaryoterna tre polymeraser som var och en består av tio underenheter eller mer., Varje eukaryotiskt polymeras kräver också en distinkt uppsättning transkriptionsfaktorer för att föra den till DNA-mallen.
RNA-polymeras I ligger i nukleolus, en specialiserad nukleär substruktur där ribosomal RNA (rRNA) transkriberas, bearbetas och monteras i ribosomer (Tabell 1). RRNA-molekylerna anses vara strukturella rna eftersom de har en cellulär roll men inte översätts till protein. RRNAs är komponenter i ribosomen och är väsentliga för översättningsprocessen. RNA polymeras i syntetiserar alla rrnas utom för 5S rRNA-molekylen., S-beteckningen gäller för Svedberg-enheter, ett icke-additivt värde som karakteriserar hastigheten vid vilken en partikelsediment under centrifugering.
| Tabell 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Eukaryotic pre-mRNAs genomgår omfattande behandling efter transkription men före översättning (Figur 1). För tydlighetens skull kommer denna moduls diskussion om transkription och översättning i eukaryoter att använda termen ”mRNAs” för att beskriva endast de mogna, bearbetade molekyler som är redo att översättas. RNA polymeras II är ansvarig för att transkribera den överväldigande majoriteten av eukaryota gener.

Figur 1. Eukaryotic mRNA innehåller introner som måste skarvas ut. En 5′ cap och 3 ’ poly-a svans tillsätts också.,
RNA polymeras III ligger också i kärnan. Denna polymeras transkriberar en mängd strukturella RNA som inkluderar 5S pre-rRNA, överföring pre-RNAs (pre-tRNAs) och små nukleära pre-RNAs. TRNAs har en kritisk roll i översättningen; de fungerar som adaptermolekyler mellan mRNA-mallen och den växande polypeptidkedjan. Små kärnkraft rna har en mängd olika funktioner, inklusive ”Splitsning” pre-mRNAs och reglera transkriptionsfaktorer.,
en forskare som karakteriserar en ny gen kan bestämma vilken polymeras som transkriberar den genom att testa om genen uttrycks i närvaro av ett visst svampgift, α-amanitin (Tabell 1). Intressant, α-amanitin produceras av Amanita phalloides, Död Cap svamp, påverkar de tre polymeraser mycket annorlunda. RNA polymeras I är helt okänslig för α-amanitin, vilket innebär att polymeras kan transkribera DNA in vitro i närvaro av detta gift. Däremot är RNA-polymeras II extremt känslig för α-amanitin, och RNA-polymeras III är måttligt känslig., Att veta transkribera polymeras kan leda en forskare till den allmänna funktionen hos genen som studeras. Eftersom RNA polymerase II transkriberar de allra flesta gener kommer vi att fokusera på detta polymeras i våra efterföljande diskussioner om eukaryotiska transkriptionsfaktorer och promotorer.
RNA Polymerase II promotorer och transkriptionsfaktorer
eukaryota promotorer är mycket större och mer invecklade än prokaryota promotorer. Båda har emellertid en sekvens som liknar -10-sekvensen av prokaryoter., I eukaryoter kallas denna sekvens Tata-rutan och har konsensussekvensen tataaa på kodningssträngen. Den ligger vid -25 till -35 baser i förhållande till initieringsstället (+1) (Figur 2). Denna sekvens är inte identisk med E. coli -10-rutan, men den bevarar a–t-rika elementet. Termostabiliteten hos A-T-bindningar är låg och detta hjälper DNA-mallen att lokalt varva ner i förberedelse för transkription.,
istället för den enkla σ-faktorn som hjälper till att binda prokaryotiskt RNA-polymeras till dess promotor, samlar eukaryoter ett komplex av transkriptionsfaktorer som krävs för att rekrytera RNA-polymeras II till en proteinkodningsgen. Transkriptionsfaktorer som binder till promotorn kallas basala transkriptionsfaktorer. Dessa basala faktorer kallas alla TFII (för transkriptionsfaktor/polymeras II) plus en extra bokstav (A-J). Kärnkomplexet är TFIID, som innehåller ett TATA-bindande protein (TBP)., De andra transkriptionsfaktorerna faller systematiskt på DNA-mallen, där var och en ytterligare stabiliserar Pre-initiationskomplexet och bidrar till rekryteringen av RNA-polymeras II.
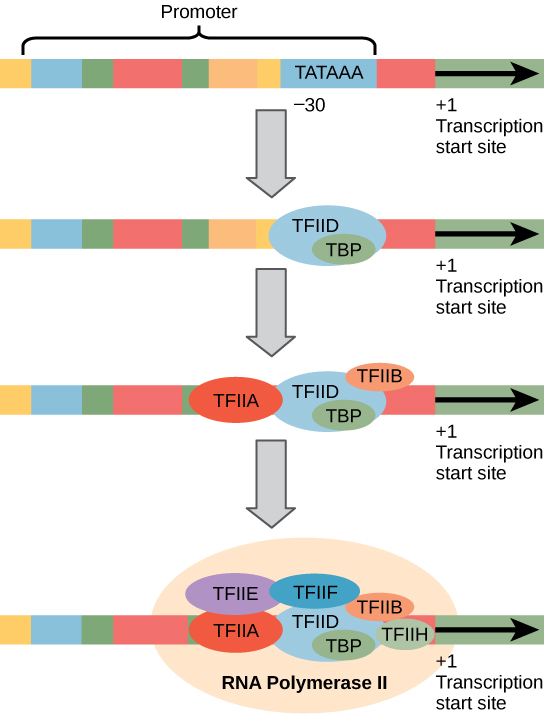
Figur 2. En generaliserad promotor av en gen transkriberad av RNA polymeras II visas. Transkriptionsfaktorer känner igen promotorn. RNA polymeras II binder sedan och bildar transkriptionsinitieringskomplexet.,
Övningsfråga
en forskare splittrar en eukaryotisk promotor framför en bakteriell gen och infogar genen i en bakteriell kromosom. Kan du förvänta dig att bakterierna transkriberar genen?
vissa eukaryota initiativtagare har också en bevarad CAAT box (GGCCAATCT) på ungefär -80. Vidare uppströms Tata-rutan kan eukaryotiska initiativtagare också innehålla en eller flera GC-rika lådor (GGCG) eller oktamer lådor (ATTTGCAT)., Dessa element binder cellulära faktorer som ökar effektiviteten av transkriptionsinitiering och identifieras ofta i mer ”aktiva” gener som ständigt uttrycks av cellen.
basala transkriptionsfaktorer är avgörande för bildandet av ett preinitiationskomplex på DNA-mallen som därefter rekryterar RNA-polymeras II för transkriptionsinitiering. Komplexiteten i eukaryotisk transkription slutar inte med polymeraser och promotorer., En armé av andra transkriptionsfaktorer, som binder till uppströmsförstärkare och ljuddämpare, hjälper också till att reglera frekvensen med vilken pre-mRNA syntetiseras från en gen. Förstärkare och ljuddämpare påverkar transkriptionens effektivitet men är inte nödvändiga för att transkriptionen ska fortsätta.
utvecklingen av promotorer
utvecklingen av gener kan vara ett välbekant koncept. Mutationer kan förekomma i gener under DNA-replikation, och resultatet kan eller inte kan vara fördelaktigt för cellen., Genom att ändra ett enzym, strukturellt protein eller någon annan faktor kan mutationsprocessen omvandla funktioner eller fysiska egenskaper. Eukaryota initiativtagare och andra genreglerande sekvenser kan dock också utvecklas. Tänk till exempel på en gen som under många generationer blir mer värdefull för cellen. Kanske kodar genen ett strukturellt protein som cellen behöver syntetisera i överflöd för en viss funktion. Om så är fallet skulle det vara fördelaktigt för cellen för den genens promotor att rekrytera transkriptionsfaktorer mer effektivt och öka genuttrycket.,
forskare som undersöker utvecklingen av promotorsekvenser har rapporterat varierande resultat. Delvis beror det på att det är svårt att dra slutsatsen exakt var en eukaryotisk promotor börjar och slutar. Vissa promotorer förekommer inom gener; andra ligger mycket långt uppströms, eller till och med nedströms, av de gener de reglerar. Men när forskare begränsade sin undersökning till human core promotorsekvenser som definierades experimentellt som sekvenser som binder preinitiationskomplexet, fann de att promotorer utvecklas ännu snabbare än proteinkodande gener.,
det är fortfarande oklart hur promotorns utveckling kan motsvara utvecklingen av människor eller andra högre organismer. Utvecklingen av en promotor för att effektivt göra mer eller mindre av en given genprodukt är dock ett spännande alternativ till utvecklingen av generna själva.
Promotorstrukturer för RNA-polymeraser I och III
processerna för att föra RNA-polymeraser I och III till DNA-mallen involverar något mindre komplexa samlingar av transkriptionsfaktorer, men det allmänna temat är detsamma.,
de konserverade promotorelementen för gener som transkriberas av polymeraser i och III skiljer sig från de som transkriberas av RNA-polymerase II. RNA-polymerase i transkriberar gener som har två GC-rika promotorsekvenser i -45 till +20-regionen. Enbart dessa sekvenser är tillräckliga för att transkriptionsinitiering ska inträffa, men initiativtagare med ytterligare sekvenser i regionen från -180 till -105 uppströms startplatsen kommer ytterligare att förbättra initieringen. Gener som transkriberas av RNA polymeras III har uppströms promotorer eller promotorer som förekommer inom generna själva.,
eukaryotisk transkription är en tätt reglerad process som kräver en mängd olika proteiner för att interagera med varandra och med DNA-strängen. Även om transkriptionsprocessen i eukaryoter innebär en större metabolisk investering än i prokaryoter, säkerställer den att cellen transkriberar exakt de pre-mRNAs som den behöver för proteinsyntes.,
Förlängning och uppsägning
Efter bildandet av preinitiationskomplexet frigörs polymeraset från de andra transkriptionsfaktorerna, och förlängning får fortsätta som det gör i prokaryoter med polymerasen som syntetiserar pre-mRNA i 5′ till 3′ – riktningen. Som diskuterats tidigare transkriberar RNA polymeras II den största andelen eukaryota gener, så det här avsnittet kommer att fokusera på hur detta polymeras åstadkommer förlängning och uppsägning.,
även om den enzymatiska töjningsprocessen i huvudsak är densamma i eukaryoter och prokaryoter, är DNA-mallen mer komplex. När eukaryota celler inte delar sig, finns deras gener som en diffus massa av DNA och proteiner som kallas kromatin. DNA: t är tätt förpackat runt laddade histonproteiner med upprepade intervall. Dessa DNA-histonkomplex, kollektivt kallade nukleosomer, är regelbundet åtskilda och inkluderar 146 nukleotider av DNA lindade runt åtta histoner som tråd runt en spole.,
för att polynukleotidsyntesen ska inträffa måste transkriptionsmaskineriet flytta histoner ur vägen varje gång det möter en nukleosom. Detta uppnås genom ett speciellt proteinkomplex som kallas FACT, som står för ”underlättar kromatin transkription.”Detta komplex drar histoner bort från DNA-mallen som polymeras rör sig längs den. När pre-mRNA syntetiseras ersätter det faktum komplexet histonerna för att återskapa nukleosomerna.
avslutandet av transkriptionen är annorlunda för de olika polymeraserna., Till skillnad från i prokaryoter sker förlängning av RNA-polymeras II i eukaryoter 1,000-2,000 nukleotider bortom slutet av genen som transkriberas. Denna pre-mRNA svans avlägsnas därefter genom klyvning under mRNA bearbetning. Å andra sidan kräver RNA-polymeraser i och III termineringssignaler. Gener transkriberade av RNA polymeras I innehåller en specifik 18-nukleotidsekvens som känns igen av ett termineringsprotein. Processen för uppsägning i RNA-polymeras III innefattar en mRNA-hårnål som liknar rho-oberoende uppsägning av transkription i prokaryoter.,
prova det
bidra!
förbättra denna sida mer
- h Liang et al.”Snabb evolution av core projektansvariga primat genom,” Molecular Biology and Evolution 25 (2008): 1239-44.















