gränser i Växtvetenskap
introduktion
korrekt progression av fruktmognad är avgörande för att uppnå både optimal fruktkvalitet och lång hållbarhet, viktiga egenskaper som bestämmer prismarknaderna och slutliga vinster. Mogning är emellertid en komplex process där många faktorer är involverade, inklusive hormonell kontroll, som reglerar biokemiska och fysiologiska förändringar som ger slutliga organoleptiska och näringsrika fruktegenskaper., Traditionellt har köttiga frukter klassificerats enligt mognadsprocessen i klimakteriella och icke-klimakteriella frukter. Klimakteriska frukter som tomater eller bananer kräver en ökning av andningsfrekvensen och etylenproduktionen för att frigöra mognadsprocessen i ett autokatalytiskt svar (Barry och Giovannoni, 2007; Liu et al., 2015)., Antagonistiskt uppvisar icke-klimakteriella frukter som citrusfrukter eller druvor inte en burst varken i eten eller andning före mogningsstart, och i dessa frukter kontrolleras mogning huvudsakligen av progressiv ackumulering av fytohormonabskisinsyran (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Nya studier förstärker dock tanken att mognadsprocessen inte bara styrs av produktionen av en fytohormon utan snarare verkar vara resultatet av en kontrollerad hormonbalans (Symons et al., 2012; Teribia m.fl., 2016; Li et al., 2019).,
auxiner, salicylsyra (SA) och melatonin är fytohormoner som är involverade i signalering och reglering av många viktiga processer i växter. Auxiner har beskrivits allmänt som tillväxt-och utvecklingsregulatorer med flera funktioner i växter (se Taylor-Teeples et al., 2016); SA utlöser försvarsrespons mot biotrofa och hemi-biotrofa patogener (Loake and Grant, 2007) samt har en viktig roll under abiotisk stress (Dong et al., 2014; Wani m.fl., 2017), blomning och cell cykel kontroll (Carswell et al. 1989; Eberhard m.fl., Och melatonin har inte bara visat sig ha auxin-liknande funktioner (Chen et al., 2009; Zuo m.fl., 2014; Wen et al., 2016) men det har också föreslagits att agera som en potentiell antioxidant i vissa växter (Arnao och Hernández-Ruiz, 2015) och en regulator av anläggningen svar att patogener (Chen et al., 2018). Intressant är att dessa tre hormoner delar en gemensam prekursor-chorismate -, vilket innebär att ett metaboliskt korssamtal uppstår mellan dem, och ett antal gener måste regleras fint för att avleda chorismate metabolism mot dessa föreningar., Under de senaste åren har flera recensioner tagit upp biosyntesen och rollen som auxin (Paul et al., 2012), SA (Asghari och Aghdam, 2010) och melatonin (Feng et al., Och med 2014. Arnao och Hernández-Ruiz, 2018) i frukt. Men deras biosyntes och funktioner har mestadels beskrivits separat. Här kommer vi att diskutera de gemensamma och differentiella aspekterna av biosyntesen, metabolismen och funktionen av auxin, SA och melatonin i tillväxten och mogningen av klimakteriella och icke-klimakteriella frukter., En betoning kommer att läggas på metabolisk avledning viktiga punkter i deras biosyntes från chorismate, den regulatoriska roll under mognaden av klimakteriska och icke-klimakteriska frukter, och deras roll i moduleringen av biosyntesen av bioaktiva föreningar, som till stor del bestämmer fruktkvalitet.
Chorismate-Derived Phytohormoner
Chorismate är slutprodukten av shikimate pathway och spelar en nyckelroll i biosyntesen av fytohormoner (Figur 1)., Chorismate ger upphov till aromatiska aminosyror, inklusive tryptofan, och genom flera reaktioner, inklusive omvandlingen av tryptofan till indol-3-pyruvsyra av tryptofanaminotransferas (TA); det kan generera auxin indol-3-ättiksyra (IAA). TA har föreslagits som ett universellt nyckelenzym till IAA biosyntes inte bara i vegetativa organ (Enríquez-Valencia et al., 2018) men också i utvecklingen av reproduktionsorgan (Reyes-Olalde et al., 2017), inklusive frukttillväxt och mogning (Estrada-Johnson et al., 2017)., Däremot, när isochorismate synthase (ICS) aktiveras, kan chorismate omvandlas till isochorismate och den senare kan omvandlas till SA. Trots det faktum att denna väg först identifierades i bakterier, är den också för närvarande väl etablerad i växter (Wildermuth et al., 2001; Uppalapati m.fl., 2007; Catinot et al., 2008; Garcion m.fl., 2008; Abreu och Munné-Bosch, 2009). Slutligen kan tryptofan omvandlas till tryptamin genom enzymet tryptofandekarboxylas (TDC)., Även om detta är en annan av de flera vägarna för IAA biosyntes (tryptamin förvandlas till indol-3-acetaldehyd och senare i IAA), har TDC också föreslagits vara det första hastighetsbegränsande enzymet i melatoninvägen (Kang et al., 2007). TDC först identifierades i familjen Apocynaceae (De Luca et al., 1989) som senare beskrivs i ett antal anläggningssystem (Byeon et al., 2012, 2014; Zhao et al., 2013; Wei m.fl., 2018)., Efter dekarboxylering omvandlas tryptamin till serotonin genom tryptamin-5-hydroxylas och sedan omvandlas serotonin till N-acetyl-serotonin för att slutligen ge melatonin i cytosolen (Figur 1).
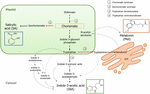
Figur 1. Biosyntes av indol-3-ättiksyra (IAA), salicylsyra (SA), och melatonin (Mel) från chorismate. Alla tre föreningarna delar en gemensam slutlig prekursor (chorismate, kvadrerad i orange) från shikimate pathway., Stora avledningspunkter indikeras med röda pilar och motsvarande enzymer numreras i en nedstigningsdivergensordning från shikimate. Djärva streckade pilar indikerar mer än ett steg mellan föreningar. Tunna streckade pilar indikerar vägar som fortfarande inte är väldefinierade.
key-diverting enzymer som leder till syntesen av chorismate-pathway hormoner har nyligen identifierats i köttiga frukter (Figur 1). ICS, vilket uttryck har visat sig vara avgörande för biotisk stresstolerans (Garcion et al.,, 2008; Zeng och He, 2010), har identifierats i tomater och äpplen. Zhu et al. (2016) rapporterade överuttrycket av ICS1 (isochorismate synthase 1) under kalla lagringsförhållanden i tomat, medan Zhang et al. (2017) fann aktivering av en transkriptionsfaktor som är involverad i den patogenrelaterade sa-signaleringsvägen som inducerar ICS-uttryck i äpplen. Förbättrad TDC aktivitet har rapporterats förekomma i omogna peppar frukt vid infektion av patogener genom ökad TDC1 och TDC2 uttryck (Park et al., 2013) – och i tillväxtfasen i mulberry frukt (Wang et al., 2016)., Slutligen har TA identifierats i grapevine, både vid pre – och véraison (Böttcher et al. 2013; för Gouthu och Deluc, 2015) samt under strawberry mognad (Estrada-Johnson et al., 2017). Det är dock anmärkningsvärt att de gener och enzymer som beskrivs i klimakteriska frukter (dvs ICS) inte har identifierats i icke-klimakteriska frukter (dvs TDC och TA) och vice versa; därför är ytterligare studier absolut nödvändiga för att fylla dessa kunskapsluckor och bättre förstå hur dessa avledningspunkter regleras gemensamt under fruktmognad.,
Roll Chorismate-Derived fytohormoner i klimakteriska och icke-klimakteriska frukter
Unraveling mekanismerna för fruktutveckling har varit en av de största utmaningarna i den senaste agronomiforskning för dess ekonomiska konsekvenser. I detta sammanhang har fytohormoner påpekats som ansvariga förare av fruktmognad, speciellt eten och ABA i klimakteriella respektive icke-klimakteriella frukter. Men att dessa fytohormoner kunde reglera fruktutvecklingen ensam visade sig snart vara alldeles för enkelt., Efter omfattande forskning och med förbättring av analytisk kemi och molekylära tekniker har flera andra hormoner bekräftats som potentiella regulatorer för fruktutveckling och mogning, inklusive chorismate-härledda fytohormoner.
auxiner Cross-Talk med andra hormoner under frukt Set, tillväxt och mognad
auxiner är en grupp av växthormoner som spelar en viktig roll i fruktutveckling, både utövar sitt eget inflytande och modulerande uttryck för andra fytohormoner., Endogena innehållet i IAA är särskilt hög vid frukt set och under inledande tillväxtutvecklingsstadier, varefter IAA-mängder tenderar att minska före mognande start, både i klimakteriet (Zaharah et al., 2012) och icke-klimakteriska frukter (Symons et al., 2012; Teribia m.fl., 2016), med tydligen några undantag, som persikor (Tatsuki et al., 2013) och några plommon sorter (El-Sharkawy et al., 2014; figur 2a). Det har visats att IAA är involverad i frukt set initiering i kombination med gibberellins (Mezzetti et al., 2004; Serrani m.fl., 2010; Bermejo et al., 2018; Hu et al.,, 2018). Försämring av IAA biosyntes eller signalering leder i allmänhet till frukt parthenocarpy, även om det också kan leda till onormal mognad i vissa frukter (Wang et al., 2005; Liu, J. et al., År 2018. Reig et al., 2018). Hög halt av IAA i de inledande stadierna av frukt utveckling främja frukttillväxt på grund av auxin implikation i celldelning i kombination med cytokininer och i kontrollen av cell expansion i kombination med gibberellins (Liao et al., 2018)., Under denna period tillåter hormonell crosstalk mellan auxins och gibberellins dessutom normal fruktformning i en finjusterad reglering medierad av Auxin Responsfaktorer (ARFs; Liao et al., År 2018. Liu S. et al., 2018).
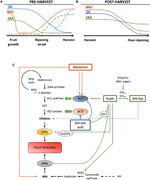
Figur 2. Roll IAA, SA och Mel under utvecklingen av klimakteriella och icke-klimakteriella frukter. Modell som sammanfattar interaktionerna mellan IAA, SA och Mel under mognaden av klimakteriella och icke-klimakteriella frukter under a) före och B) efter skörd., Streckade linjer indikerar alternativ dynamik av fytohormon innehåll i vissa frukter (se text för diskussion). C) översikt över interaktionen mellan IAA, SA och Mel med etylen och abscisinsyra (ABA) biosyntes i klimakteriella och icke-klimakteriella frukter. Auxin är en positiv regulator av etylenbiosyntes genom aktivering av ACC-syntasgener (ACS) och inducerar uttrycket av Etylenresponsiva faktorer (ERFs). Auxin undertrycker också ABA-produktionen av 9-cis-epoxikarotenoid dioxygenas (NCED) förtryck och hämmar ABA-responsiva stressgener (ARS) av ARF2-proteinet., Endogent IAA-innehåll kan minskas med GH3-proteiner, vars syntes främjas av eten, ABA och sockerarter. Diskontinuerlig linje indikerar IAA-Asp: s möjliga roll som en mognadsfaktor som förbättrar eten i vissa frukter. Melatonin ökar ABA-och etylenproduktionen, även om ACS kan hämmas beroende på melatonininnehåll. SA verkar som en hämmare av etylen som undertrycker ACC-oxidas (Aco) gener, även om det kan främja uttrycket av ACS., Mta, 50-metyltioadenosin; s-AdoMet, S-adenosyl-metionin; SAMSYNTAS, S-adenosyl-metioninsyntas; ACC, 1-aminocyklopropan-1-karboxylsyra; IPP, isopentenyldifosfat.
minskningar av endogent IAA-innehåll har rapporterats inträffa före mogning i flera frukter. Dessa minskningar har relaterats till IAA-konjugering med asparaginsyra (IAA-Asp) av IAA-amido synthetaser, GH3-1 och GH3-2 (Figur 2C)., GH3 – 1 och GH3-2 visade faktiskt högre uttryck under tidig fruktutveckling och framför allt under mognadsinitiering både i klimakteriska frukter, såsom tomater (Sravankumar et al., 2018) och äpplen (Onik et al., 2018), liksom i icke-klimakteriska frukter, som druvbär (Böttcher et al., 2010, 2011) och hallon (Bernales et al., 2019)., Intressant, druvbär visade förbättrad GH3 – 1 uttryck efter ABA och etefon ansökan, vilket skulle kunna förklara inblandning av eten i kontrollen av IAA innehåll efter starten av mognad, även i icke-klimakteriska frukter (Böttcher et al., 2010). Faktum är att flera studier belyser den snäva interaktionen mellan auxiner och eten i fruktmognad, med ett ömsesidigt inflytande mellan dem (Tadiello et al., 2016a; Busatto et al., 2017)., För klimakteriska frukter är ökat innehåll av IAA nödvändigt för att aktivera uttryck av ACC-syntasgener (ACS), som kodar för ACC-syntas, vilket i sin tur leder till etylenproduktion som utlöser mognadsprocessen (figur 2C; Tatsuki et al., 2013). Frukt som tomater (Li et al., 2016) eller bananer (Choudhury et al.,, 2008), som vanligtvis upplever minskat IAA-innehåll före mognadens början, visar en mognadsfördröjning när IAA eller analoger appliceras exogent, medan frukter som persikor, vars IAA-innehåll ökar gradvis tills de är helt mogna, visar accelererad mogning när auxiner appliceras (Tadiello et al., 2016b). Därför måste en snäv, komplex och differentiell reglering av auxin-etyleninteraktioner existera i olika frukter under preharvest mogning., Dessutom minskar IAA-innehållet före mogning i vissa icke-klimakteriella frukter för att avtrycka 9-cis-epoxikarotenoid dioxygenas (NCED) och starta ABA-syntes (figur 2C; Jia et al., 2016). Intressant är att denna process också medieras av IAA-konjugation genom förbättrad GH3-1-aktivitet (Böttcher et al., 2010). Ändå, Estrada-Johnson et al. (2017) visade att även om IAA-innehållet minskar under jordgubbsmognad induceras uttryck av FaAux/IAA och FaARF-genfamiljer i röda behållare, vilket tyder på att auxin-signalering är inblandad i fullt mogna frukter.,
Under efter skörd förblir auxininnehållet vanligtvis oföränderligt eller tenderar att minska på grund av oxidativa processer som kan ge upphov till små men progressiva minskningar av det endogena innehållet i IAA (Figur 2B). Auxin behandlingar efter skördefördröjning övermognad i vissa frukter (Chen et al., 2016; Moro et al., 2017) och öka innehållet i vissa organiska syror, upprätthålla fruktsyra (Li et al., 2017), vilket tyder på att auxin också spelar en viktig roll i kontrollen av fruktmognad under efterskörden.,
flera roller av salicylsyra under fruktutveckling och mogning
salicylsyra är en annan chorismate-härledd fytohormon som mestadels har relaterats till dess skyddande effekt under biotisk stress för att kontrollera preharvest och efter skörd förluster härrörande från patogen fruktinfektion (Babalar et al., 2007; Cao m.fl., 2013). I allmänhet är endogena innehållet i free SA högre i början av fruktutvecklingen och minskar sedan gradvis (Oikawa et al., 2015). Lu et al. (2015) rapporterade en sekundär ökning av SA under den andra tillväxtfasen av persika frukter., Exogen applicering av SA i bananer försenade mognadsprocessen som minskar andningsbrottet och förhållandet mellan massa och skal, liksom minskande aktivitet av enzymer relaterade till cellväggsnedbrytning och antioxidantsystem (Srivastava och Dwivedi, 2000). Dessutom, behandling av söta körsbärsträd med salicylsyra och acetylsalicylsyra (ASA) förbättrad vikt, fasthet och färg av körsbär i kommersiellt skede (Giménez et al., 2014). Exogen applicering av SA eller metyl salicylat (MeSA) under efter skörd, fördröjd övermognad i frukter som kiwi (Zhang et al.,, 2003), och sötkörsbär (Valero et al., 2011). Dessutom behandlingar med salicylsyra eller ASA lindras kylning skada av granatäpplen (Sayyari et al., 2011), tomater (Aghdam et al., 2014), och avokado (Glowacz et al., 2017). SA interagerar även med eten genom hämning av ACC oxidas (ACO) uttryck (Shi och Zhang, 2012). Förvånansvärt finns också ökat innehåll av ACS-uttryck i päron efter sa-applicering på ett dosreglerat sätt (Shi et al. 2013; för Shi och Zhang, 2014). Därför är regleringen av etenproduktion beroende av SA fint reglerad (figur 2C).,
Melatonin: en framväxande Regulator för kontroll av fruktutveckling
Melatonin är också ett växthormon från den chorismate-härledda vägen som har fått stor uppmärksamhet de senaste åren (Hernández-Ruiz et al., 2005; Arnao och Hernández-Ruiz, 2018; Sharif et al., 2018). Nyligen har melatonin påpekats som en viktig faktor för fruktuppsättning, på ett liknande sätt som IAA-effekter (figur 2a; Liu S. et al., 2018). Dessutom har cirkadisk produktion av denna fytohormon observerats (Kolář et al., 1997)., Vid mogning av träd visade druvor behandlade med melatonin högre synkronicitet och ökad vikt när de mognar fullt ut (Meng et al., 2015). Dessutom en annan nyligen genomförd studie av Xu et al. (2018) beskrev att melatonin kunde främja druvbärmognad genom dess interaktion med ABA och etylen, liksom väteperoxid. Det har visat sig att melatonin ger kylningstolerans under fruktkylförvaring, vilket verkar vara mest relaterat till dess antioxidantaktivitet (Cao et al., År 2018. Aghdam et al., 2019)., Under efter skörd visade tomater ökat etylenutsläpp efter applicering av melatonin vid 50 µM (Sun et al., 2015), i motsats till päron, där exogena tillämpning av melatonin vid 100 µM hämmade etylenproduktion (Zhai et al., 2018), eller bananfrukter, där melatonin vid 50-200 µM hämmade uttrycket av ACS och Aco (figur 2C; hu et al., 2017). Även om olika effekter av melatonin ansökan kan vara relaterade till en doseringseffekt, ytterligare forskning behövs för att bättre förstå rollen av melatonin i kontrollen av frukt utveckling.,
Cross-Talk Of Chorismate-Derived Phytohormoner in the Control of Fruit Development
hormonell crosstalk mellan chorismate-derived phytohormoner har visat sig förekomma under fruktutveckling och mogning. Breitel et al. (2016) fann att auxiner kan interagera med SA genom ARF2 (figur 2c) och att överuttryck av arf2 i tomat resulterade i lägre innehåll av SA och en signifikant mognadsfördröjning., Detta indikerar att auxiner kan begränsa SA-produktionen under fruktutveckling, vilket kan förklara en avvägning mellan frukttillväxt (medierad av auxin) och aktivering av biotiska försvar (medierad av SA). Dessutom, exogena tillämpning av SA på papaya frukter resulterade i en förändrad uttryck av flera IAA gener, några av dem är nedreglerade medan andra uppreglerade (Liu et al., 2017)., I det här fallet var resultaten ofullständiga av vad som verkligen uppstår endogent i det putativa korssamtalet mellan auxin och SA under fruktmognad och ytterligare forskning krävs i denna och andra klimakteriella frukter. I vilket fall som helst är det uppenbart att IAA och SA är nära besläktade, inte bara metaboliskt utan också funktionellt. Slutligen har flera studier rapporterat påverkan av melatonin på IAA och sa biosyntes och / eller signalering (granskad i Arnao och Hernández-Ruiz, 2018), även om vi vet att ingen av dessa studier utfördes varken i klimakterisk eller i icke-klimakterisk frukt.,
modulering av bioaktiva föreningar biosyntes av Chorismate-Derived fytohormoner
bioaktiva föreningar (inklusive fenolföreningar, isoprenoider och antioxidant vitaminer) har inte bara undersökts i stor utsträckning som ansvarig för specifika organoleptiska egenskaper hos livsmedel, men också för deras skyddande effekter i humana celler mot oxidativa processer vid utveckling av neurodegenerativa och kardiovaskulära sjukdomar och viss typ av cancer (Liu et al., 1999; Müller et al., 2010; Stör och Ronnenberg, 2010; Björkman et al., 2011)., Fenolföreningar bidrar avsevärt till att ge specifika smaker, såsom tanniner-ansvarig för bitterhet eller astringency smak av vissa frukter och färger, såsom anthocyaninpigment-ansvarig för röda, blå och lila fruktfärger (Croteau et al., 2000). Betydelsen av fenolföreningar som livsmedel har förknippats med en skyddande effekt mot oxidativa processer i förhållande till hjärt-och centrala nervsystemet samt en minskad risk för cancer i mag-tarmkanalen (Björkman et al., 2011)., Karotenoider, tetraterpener som tillhör isoprenoider, spelar en roll i skyddet mot fotooxidativa processer och som organiska pigment; de är ansvariga för den orangegula färgen på frukter (tapiero et al., 2004). Dietary karotenoider antas ge hälsofördelar för att minska risken för ögonsjukdom och vissa typer av cancer på grund av deras roll som antioxidanter (Johnson, 2002). Vitaminer C och E, inklusive askorbinsyra och tokoferoler, är väsentliga näringsmässiga kvalitetsfaktorer i frukter med många biologiska aktiviteter hos människor (Miret och Müller, 2017)., Med tanke på den relevanta rollen för alla dessa bioaktiva föreningar som hälsofrämjande föreningar i frukter är moduleringen av deras ackumulering av största vikt i både klimakteriella och icke-klimakteriella frukter (Vasconsuelo och Boland, 2007). De flesta av dessa bioaktiva föreningar ackumuleras på höga nivåer under fruktmognad, när mindre välsmakande gröna frukter konverseras till en näringsrikt, färgad och god frukt (Daood et al., 1996; Ranalli m.fl., 1998; Dumas m.fl., 2003; Singh et al., 2011)., Men när mognaden påbörjats kan den inte stoppas och leder i allmänhet till övermognad som negativt påverkar fruktkvaliteten (Kumar et al., 2014). Därför är minimering av fruktförstöring efter skörden samtidigt som man får högt innehåll av bioaktiva föreningar fortfarande en av de största utmaningarna att lösa.
Auxin har identifierats som en viktig regulator för karotenoidbiosyntes under mogning av klimakteriella tomatfrukter. Mognad i tomat är förknippad med nedbrytningen av klorofyll och förskjutningen av xantofyller till karotener (β-karoten och lykopen) (Fraser et al., 1994)., IAA verkar dröjsmål tomat mognad genom att undertrycka eten och flera uppströms karotenoid avskrifter inklusive Psy, Ziso, Pds, Critiso liksom Chlases 1-3 och, å andra sidan, att främja ansamling av β-Lyc1 och Crtr-β1 avskrifter, vilket resulterar i högre halter av xantofyller och klorofyll a (Su et al., 2015). Dessutom verkar i vissa icke-klimakteriska frukter som körsbär och druvbär auxin också modulera antocyaninbiosyntes och därigenom styra fruktmognadsprocesser. Teribia et al., (2016) rapporterade en negativ korrelation mellan IAA och antocyanininnehåll vilket indikerar att antocyaninackumulering börjar när IAA-innehållet minskar. Dessutom fördröjde appliceringen av den syntetiska auxinliknande föreningen bensotiazol-2-oxyacetic acid (BTOA) uppregleringen av gener som främjar enzymer av antocyaninbiosyntesen såsom chalkonsyntas och UDP-glukos:flavonoid 3-o-glukosyltransferas i druvbär (Davies et al., 1997).,
behandling före och efter skörd med salicylater, inklusive SA och dess derivat Asa och MeSA, har rapporterats reglera ackumulering av bioaktiva föreningar som leder till förbättrad antioxidantaktivitet i både klimakteriella och icke-klimakteriella frukter. Flera frukter som plommon, körsbär och aprikoser är kända för att ha ett kort liv med en snabb försämring av kvaliteten efter skörden. Därför finns det en ständig sökning av behandlingar som förbättrar och upprätthåller fruktkvalitet, och särskilt innehållet i bioaktiva föreningar med fördelaktiga hälsoeffekter., Behandlingar av plommon med SA, Asa och MeSA resulterade i signifikant högre innehåll av askorbinsyra, antocyaniner och fenolföreningar både vid skörd och efter långvarig kylförvaring (40 dagar vid 4°C; Martínez-Esplá et al., 2017). Liknande resultat kunde observeras för preharvest behandlingar med SA, ASA, och MeSA i söta körsbär (Giménez et al., 2014, 2015, 2017). Vidare upprätthöll behandling efter skörd med SA, ASA eller MeSA totalt fenolinnehåll samt antocyanininnehåll under kylförvaring i granatäpple (Sayyari et al., 2011), söta körsbär (Valero et al.,, 2011), karneol körsbär (Dokhanieh et al., 2013), och aprikos (Wang et al., 2015). Dessa resultat tyder på att salicylater kan vara involverade i aktiveringen av fenylalanin ammoniaklyas, vilket är det huvudsakliga enzymet som är involverat i biosyntetiska vägen för fenolföreningar (Martínez-Esplá et al., 2017).
nyligen har melatonin visat sig reglera fruktmognad och modulera bioaktiva föreningar. Efter skörd behandlingar med melatonin i persika frukter ökade kylning tolerans genom att aktivera antioxidantsystem., Chilling skada uppstår i persika frukter under låg temperatur Lagring (mellan 2 och 7.6°C) kännetecknas av kött bryning, onormal mognad och högre känslighet för förfall (Lurie och Crisosto, 2005). Cao et al. (2018) rapporterade att persikor behandlade med melatonin visade högre transkript överflöd av askorbinsyrabiosyntesgener PpGME, PpGPP och PpGLDH vid sjunde och PpGMPH vid den 21: e och 28: e lagringsdagen jämfört med kontroll vilket resulterar i ökat askorbinsyrainnehåll., Dessutom inducerade melatonin ökningar i aktiviteter av g6pdh, SKDH och PAL, vilka är de väsentliga enzymerna för fenolföreningar biosyntes. Författarna föreslår att melatonin behandling skyddar persika frukt till en viss grad från kylning skada genom att specifikt aktivera biosyntesen av fenolföreningar (Gao et al., 2018). Dessutom avslöjade en etikettfri differentialproteomikanalys effekten av melatonin på att främja fruktmognad och antocyanin ackumulering efter skörd i tomatfrukter (Sun et al., 2016)., Författarna rapporterade att exogent melatonin ökade åtta enzymer relaterade till antocyaninvägen inklusive bland annat flavonol-3-hydroxylas, flavanon 3 beta-hydroxylas, antocyanidin syntas / leucoanthocyanidin dioxygenas och antocyanidin 3-o-glukosyltransferas. Dessutom efter skörd behandling med melatonin ökade totala fenoler och antocyaniner i jordgubbar frukter (Aghdam och ister, 2016), och fördröjd förlust av totala fenoler, flavonoider och antocyaniner i litchi frukter (Zhang et al., 2018)., Behandlingar med melatonin ökade innehållet av fenoler, antocyaniner och flavonoider i druvbär (Xu et al., 2017). Dessutom har melatonin rapporterats förbättra lykopenackumulering och etylenproduktion i tomater, vilket tyder på att melatonin kan öka innehållet av lykopen genom att påverka etenbiosyntes och signalering (Sun et al., 2015)., Etylens deltagande i melatoninininducerad reglering av bioaktiva föreningar har också observerats i druvbär, där dubbelblocksbehandling av eten visade minskade effekter av melatonin på polyfenolinnehåll (Xu et al., 2017).
slutsats
man drar slutsatsen att fytohormoner som härrör från choriskompis, inklusive auxin, SA och melatonin, inte bara delar en gemensam föregångare utan spelar en viktig roll vid reglering av frukttillväxt och mogning., En metabolisk och funktionell korssamtal mellan dem och med andra fytohormoner uppträder på ett spatiotemporalt sätt för att fint reglera utvecklingen av klimakteriella och icke-klimakteriella frukter. Skillnaderna i dynamiken inom klimakteriella och icke-klimakteriella frukter bevisar att svaret från chorismate-härledda hormoner inte är universellt men ganska starkt artspecifikt. Dessutom modulerar chorismate-härledda fytohormoner också ackumuleringen av bioaktiva föreningar, vilket påverkar fruktkvaliteten., Ytterligare forskning behövs dock för att bättre förstå hur (i) andra hormoner, såsom etylen och ABA, modulerar deras biosyntes som studerar viktiga metaboliska avledningspunkter, (ii) de interagerar på funktionella och molekylära nivåer, och (iii) de modulerar bioaktiva föreningar biosyntes och påverkar följaktligen inte bara frukttillväxt och mogning utan också kvaliteten på klimakteriella och icke-klimakteriella frukter.
författare bidrag
SM-b tänkt och utformat översynen med hjälp av MP-L. MP-l, PM, MM, och SM-b skrev manuskriptet., MP-l beredda figurer med hjälp av PM. Alla författare bidrog till diskussionen om idéer, reviderade och godkände det slutliga manuskriptet.
finansiering
detta arbete finansierades av en ICREA Academia award ges till SM-B av den katalanska regeringen.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Böttcher, C., Boss, P. K., och Davies, C. (2011)., Acylsubstratpreferenser för en IAA-amido-syntetas står för variationer i druv (Vitis vinifera L.) bärmognad orsakad av olika auxinföreningar som indikerar vikten av auxin-konjugering vid växtutveckling. J. Exp. Bot. 62, 4267–4280. doi: 10.1093/jxb/err134
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., och Sivakumar, D. (2017)., Upprätthålla postharvest kvaliteten på kall lagrade ”Hass” avokado genom att ändra fettsyrorna innehåll och sammansättning med användning av naturliga flyktiga föreningar – metyl jasmonat och metylsalicylat. J. Sci. Mat. Agric. 97, 5186–5193. doi: 10.1002/jsfa.8400
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Zeng, W., och Han, S. Y. (2010). En framträdande roll i flagellin receptor FLAGELLIN-SENSING2 medla stomatala svar på Pseudomonas syringae pv tomat DC3000 i Arabidopsis. Växtfysiol. 153, 1188–1198. doi: 10.1104/s. 110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















