Biologi for Hovedfag I
læringsutbytte
- Beskrive rollen av RNA polymerase
Dette avsnittet vil utvide på den spesifikke rollen som RNA polymerases under transkripsjon. Les videre for å lære rollen som RNA polymerases på hvert trinn av transkripsjon.,
Initiering av Transkripsjon
i Motsetning til den prokaryotic polymerase som kan binde seg til DNA-mal på sin egen, eukaryotes krever flere andre proteiner, som kalles transkripsjon faktorer, først binder seg til arrangøren region og deretter hjelpe rekruttere de riktige polymerase.
De Tre Eukaryote RNA Polymerases
funksjoner av eukaryote mRNA-syntese er markant mer komplekse de av prokaryotes. I stedet for en enkelt polymerase bestående av fem underenhetene, den eukaryotes har tre polymerases som hver består av 10 underenhetene eller mer., Hver eukaryote polymerase også krever et spesifikt sett av transkripsjon faktorer for å få det til DNA-templat.
RNA-polymerase jeg er forretningsvennlig og ligger i nucleolus, en spesialisert kjernefysiske underlaget der ribosom-RNA (rRNA) er transkribert, bearbeidet og satt sammen til ribosomes (Tabell 1). RRNA-molekylene er vurdert strukturelle RNAs fordi de har en mobil rolle, men er ikke oversatt til proteiner. Den rRNAs er komponenter i ribosomet og er avgjørende for prosessen med oversettelse. RNA-polymerase jeg syntetiserer alle rRNAs bortsett fra 5S rRNA-molekylene., «S» – betegnelsen som gjelder for «Svedberg» enheter, en nonadditive verdi som karakteriserer den hastigheten som en partikkel sedimenter under sentrifugering.
| Tabell 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Eukaryote pre-mrnaer gjennomgår omfattende behandling etter transkripsjon men før oversettelse (Figur 1). For klarhet, denne modulen er diskusjon av transkripsjon og oversettelse i eukaryotes vil bruke begrepet «mrnaer» for å beskrive kun modne, behandlet molekyler som er klar til å bli oversatt. RNA polymerase II er ansvarlig for å transkribere det overveldende flertallet av eukaryote gener.

Figur 1. Eukaryote mRNA inneholder introns som må være splittet ut. 5′ cap og 3′ poly-A hale er også lagt til.,
– >
RNA-polymerase III er også plassert i kjernen. Dette polymerase transcribes en rekke strukturelle RNAs som inkluderer 5S pre-rRNA, overføre pre-RNAs (pre-tRNAs), og liten kjernefysisk pre-RNAs. Den tRNAs har en avgjørende rolle i oversettelse; de tjener som adapter molekyler mellom mRNA mal og voksende polypeptid-kjeden. Liten kjernefysisk RNAs har en rekke funksjoner, inkludert «skjøting» pre-mrnaer og regulering av transkripsjon faktorer.,
En vitenskapsmann som karakteriserer et nytt gen kan bestemme hvilke polymerase transcribes det ved å teste om genet er uttrykt i nærvær av en bestemt sopp gift, α-amanitin (Tabell 1). Interessant, α-amanitin produsert av Amanita phalloides, Død Cap sopp, påvirker de tre polymerases svært forskjellig. RNA-polymerase jeg er helt upåvirkelig av α-amanitin, noe som betyr at polymerase kan transkribere DNA in vitro i nærvær av denne giften. I kontrast, RNA polymerase II er ekstremt følsom for α-amanitin, og RNA-polymerase III er moderat følsomme., Å vite transkribere polymerase kan ane en forsker til den generelle funksjonen til genet som blir studert. Fordi RNA polymerase II transcribes det store flertallet av gener, vil vi fokusere på dette polymerase i våre diskusjoner om eukaryote transkripsjonfaktorer og arrangører.
RNA Polymerase II Arrangører og transkripsjonfaktorer
Eukaryote arrangører er mye større og mer innviklet enn prokaryotic arrangører. Men begge har en sekvens lik -10 sekvens av prokaryotes., I eukaryotes, denne sekvensen kalles TATA-boksen, og har konsensus sekvens TATAAA på coding strand. Den ligger ved -25 til -35 baser i forhold til initiering (+1) området (Figur 2). Denne sekvensen er ikke identiske med E. coli -10 boksen, men det sparer den På rik element. Den thermostability På obligasjoner er lav, og dette bidrar til at DNA-templat til lokalt slappe av i forberedelse for transkripsjon.,
i Stedet for den enkle σ faktor som bidrar til å binde prokaryotic RNA polymerase til arrangøren, eukaryotes sette sammen et kompleks av transkripsjonfaktorer som kreves for å rekruttere RNA polymerase II til en protein-kodende gener. Transkripsjonfaktorer som binder seg til arrangøren kalles basal transkripsjon faktorer. Disse basal faktorer er alle kalt TFII (for transkripsjonsfaktor/polymerase II) pluss en ekstra bokstav (A-J). Kjernen i komplekset er TFIID, som inkluderer en TATA-bindende protein (TBP)., Den andre transkripsjonfaktorer systematisk med å falle på plass på DNA-templat, med hver og en ytterligere stabilisering av pre-initiering komplekse, og å bidra til rekruttering av RNA polymerase II.
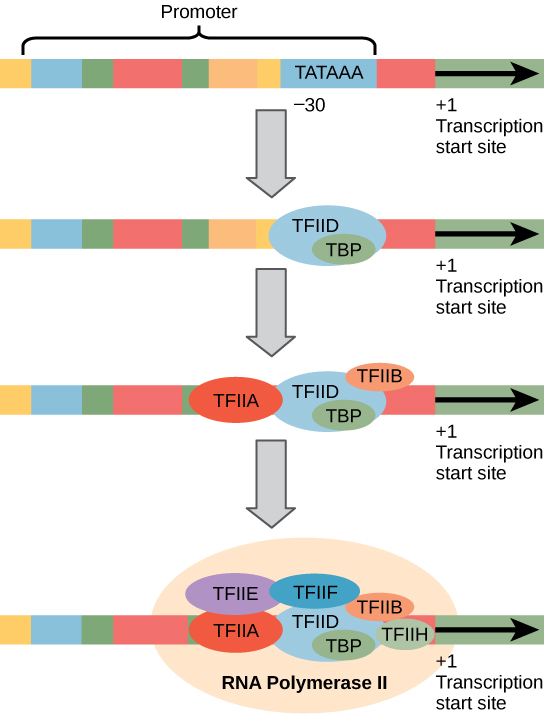
Figur 2. En generalisert arrangøren av et gen transkribert av RNA polymerase II er vist. Transkripsjonfaktorer gjenkjenne arrangøren. RNA polymerase II deretter binder seg og danner transkripsjon initiation komplekse.,
Praksis Spørsmålet
En vitenskapsmann splint en eukaryote arrangøren i front av en bakterielle genet og setter genet i en bakteriell kromosom. Ville du forvente at bakterier å transkribere genet?
Noen eukaryote organisasjonene også har en bevart CAAT-boksen (GGCCAATCT) på ca -80. Videre oppstrøms av TATA-boksen, eukaryote prosjektpartnere kan også inneholde én eller flere GC-rike bokser (GGCG) eller octamer bokser (ATTTGCAT)., Disse elementene binde cellulære faktorer som øker effektiviteten av transkripsjon initiering og er ofte identifisert i mer «aktiv» gener som stadig blir uttrykt av cellen.
Basal transkripsjon faktorer er avgjørende for dannelsen av en preinitiation kompleks på DNA-templat som senere rekrutter RNA polymerase II for transkripsjon initiering. Kompleksiteten i eukaryote transkripsjon slutter ikke med polymerases og arrangører., En hær av andre transkripsjon faktorer, som binder seg til oppstrøms enhancers og lyddempere, også bidra til å regulere frekvensen med pre-mRNA er syntetisert fra et gen. Enhancers og lyddempere påvirke effektiviteten av transkripsjon, men er ikke nødvendig for transkripsjon til å fortsette.
Utviklingen av Arrangører
evolusjon av gener kan være et kjent konsept. Mutasjoner kan oppstå i genene i DNA-replikasjon, og resultatet kan eller ikke kan være gunstig for den cellen., Ved å endre et enzym, strukturelle proteiner, eller noen annen faktor, prosessen med mutasjon kan forvandle funksjoner eller fysiske funksjoner. Imidlertid, eukaryote arrangører og andre gen regulatoriske sekvenser kan utvikle seg som godt. For eksempel bør du vurdere et gen som, over mange generasjoner, blir mer verdifulle for cellen. Kanskje genet koder et strukturelt protein som cellen trenger for å syntetisere i overflod for en bestemt funksjon. Hvis dette er tilfelle, vil det være nyttig å cellen for at genets promoter å rekruttere transkripsjonfaktorer mer effektivt og øke genuttrykk.,
Forskere å undersøke utviklingen av arrangøren sekvenser har rapportert om varierende resultater. I del, dette er fordi det er vanskelig å utlede nøyaktig hvor en eukaryote arrangøren begynner og slutter. Noen arrangører skje i løpet av gener, andre er plassert veldig langt oppstrøms, eller selv nedstrøms, gener og de er regulering. Men når forskerne begrenset deres undersøkelse for å menneskelig core promoter sekvenser som ble definert som eksperimentelt sekvenser som binder preinitiation komplekse, fant de at arrangørene utvikle seg enda raskere enn protein-kodende gener.,
Det er fortsatt uklart hvordan arrangøren utviklingen kan svarer til utviklingen av mennesker eller andre organismer høyere. Men utviklingen av en søker å effektivt gjøre mer eller mindre av et gitt gen produktet er et spennende alternativ til evolusjon av gener seg selv.
Arrangøren Strukturer for RNA Polymerases jeg og III
Den prosesser for å bringe RNA polymerases jeg og III til DNA-templat innebære litt mindre komplekse samlinger av transkripsjon faktorer, men det generelle temaet er det samme.,
bevart arrangøren elementer for gener transkribert av polymerases jeg og III er forskjellige fra de transkribert av RNA polymerase II. RNA-polymerase jeg transcribes gener som har to GC-rike arrangøren sekvenser i -45 til +20-regionen. Disse sekvensene alene er tilstrekkelig for transkripsjon initiering til å skje, men søkere med flere sekvenser i regionen fra -180 å -105 oppstrøms av initiering området vil ytterligere forsterke initiering. Gener som er transkribert av RNA polymerase III har oppstrøms søkere eller søkere som oppstår i løpet av de gener seg selv.,
Eukaryote transkripsjon er en strengt regulert prosess som krever et utvalg av proteiner for å samhandle med hverandre og med DNA-strand. Selv om prosessen med transkripsjon i eukaryotes innebærer en større metabolsk investeringer enn i prokaryotes, det sørger for at cellen transcribes nettopp pre-mrnaer at det er behov for protein syntese.,
Forlengelse og Opphør
Følgende dannelsen av preinitiation komplekse, polymerase er gitt ut fra andre transkripsjonfaktorer, og tøyelighet er lov til å fortsette som det gjør i prokaryotes med polymerase syntetisere pre-mRNA i 5′ til 3′ retning. Som diskutert tidligere, RNA polymerase II transcribes store deler av eukaryote gener, så denne delen vil fokusere på hvordan dette polymerase oppnår forlengelse og opphør.,
Selv om det enzymatisk prosess med forlengelse er i hovedsak de samme i eukaryotes og prokaryotes, DNA-malen er mer komplekse. Når eukaryote celler er ikke å dele, deres gener eksistere som en diffus masse av DNA og proteiner som kalles chromatin. DNA er tett pakket rundt belastet histone proteiner ved gjentatte intervaller. Disse DNA–histone komplekser, samlet kalt nucleosomes, er regelmessig linjeavstand og inkluderer 146 nukleotider av DNA såret rundt åtte histones som tråden rundt spolen.,
For polynucleotide syntese til å skje, transkripsjon maskiner behov for å flytte histones ut av veien hver gang den møter en nucleosome. Dette gjøres ved en spesiell protein kompleks kalt FAKTUM, som står for «forenkler chromatin transkripsjon.»Dette komplekset trekker histones bort fra DNA-templat som polymerase beveger seg langs den. Når pre-mRNA er syntetisert, det FAKTUM komplekse erstatter histones å gjenskape nucleosomes.
opphør av transkripsjon er forskjellig for de ulike polymerases., I motsetning til i prokaryotes, forlengelse av RNA polymerase II i eukaryotes foregår 1000 til 2000 nukleotider utover slutten av genet blir transkribert. Denne pre-mRNA halen er senere fjernet ved spalting i løpet av mRNA behandling. På den annen side, RNA polymerases jeg og III kreve oppsigelse signaler. Gener transkribert av RNA polymerase jeg inneholde en bestemt 18-nukleotid-sekvensen som er anerkjent av en oppsigelse protein. Prosessen med avslutning i RNA-polymerase III innebærer en mRNA hårnål lik rho-uavhengig opphør av transkripsjon i prokaryotes.,
Prøv Det
Bidra!
Forbedre dette pageLearn Mer
– >
- H Liang et al., «Raske utviklingen av kjernen arrangører i primat genomer,» Molekylær Biologi og Evolusjon 25 (2008): 1239-44. ↵















