Frontiers in Plant Science
Innledning
Riktig progresjon av frukt modning er avgjørende for å oppnå både optimal frukt kvalitet og lang holdbarhet, viktige egenskaper som bestemmer prisen markeder og endelige resultat. Imidlertid, modning er en kompleks prosess der mange faktorer som er involvert, inkludert hormonell kontroll, som regulerer biokjemiske og fysiologiske endringer som kan gi endelige organoleptiske og ernæringsmessige frukt egenskaper., Tradisjonelt, kjøttfull frukt har blitt klassifisert i henhold til det modningsprosessen i climacteric og ikke-climacteric frukt. Climacteric frukt som for eksempel tomater eller bananer krever en oppsving i åndedrett pris og etylen produksjon for å utnytte den modningsprosessen i en autocatalytic svar (Barry og Giovannoni, 2007; Liu et al., 2015)., Antagonistically, ikke-climacteric frukt som sitrus frukter eller druer ikke viser et utbrudd verken i etylen heller åndedrett før modning utbruddet, og i disse fruktene, modning er i hovedsak kontrollert av progressive opphopning av phytohormone abscisic syre (ABA; Rodrigo et al., 2006; Castellarin et al., 2011). Likevel, nye studier forsterke ideen om at modningsprosessen er ikke bare styrt av produksjonen av en phytohormone men heller tilsynelatende ut til å være resultatet av en kontrollert hormonell balanse (Symons et al., 2012; Teribia et al., 2016; Li et al., 2019).,
Auxins, salicylic acid (SA), og melatonin er phytohormoner involvert i signalering og regulering av mange viktige prosesser i planter. Auxins har vært mye beskrevet som vekst og utvikling regulatorer med flere funksjoner i planter (se Taylor-Teeples et al., 2016); SA utløser forsvar reaksjon mot biotrophic og hemi-biotrophic patogener (Loake og Grant, 2007), samt å ha en viktig rolle under abiotisk stress (Dong et al., 2014; Wani et al., 2017), blomstrende og celle-syklus kontroll (Carswell et al., 1989; Eberhard et al.,, 1989, henholdsvis), og melatonin har ikke bare blitt funnet å ha auxin-lignende funksjoner (Chen et al., 2009; Zuo et al., 2014; Wen et al., 2016), men det har også vært foreslått å fungere som en antioksidant potensial i noen planter (Arnao og Hernández-Ruiz, 2015) og en regulator av anlegget svar til patogener (Chen et al., 2018). Det er interessant at disse tre hormoner dele en felles forløper – chorismate -, og dermed en metabolsk cross-talk oppstår mellom dem, og en rekke gener må være fint regulert for å viderekoble chorismate metabolisme mot disse stoffene., I løpet av de siste årene, og flere anmeldelser har tatt for biosyntese og rollen av auxin (Paul et al., 2012), SA (Asghari og Aghdam, 2010) og melatonin (Feng et al., 2014; Arnao og Hernández-Ruiz, 2018) i frukt. Men, deres biosyntese og funksjoner har for det meste blitt beskrevet separat. Her skal vi diskutere felles og ulike aspekter ved biosyntese, metabolisme og funksjon av auxin, SA og melatonin i vekst og modning av climacteric og ikke-climacteric frukt., En hovedvekt vil bli lagt på metabolske avledning viktige poeng i sine biosyntese fra chorismate, regulerende rolle under modning av climacteric og ikke-climacteric frukt, og deres rolle i modulering av biosyntese av bioaktive forbindelser, som i stor grad bestemmer frukt kvalitet.
Chorismate-Avledet Phytohormoner
Chorismate er det endelige produktet av shikimate veien og spiller en viktig rolle i biosyntesen av phytohormoner (Figur 1)., Chorismate gir opphav til aromatiske aminosyrer, inkludert tryptofan, og gjennom flere reaksjoner, inkludert omdanningen av tryptofan til indole-3-pyruvic syre av tryptofan aminotransferase (TA); det kan generere auxin indole-3-eddiksyre (IAA). TA har vært foreslått som en universell viktig enzym for å IAA biosyntese ikke bare i vegetative organer (Enríquez-Valencia et al., 2018), men også i utviklingen av reproduktive organer (Reyes-Olalde et al., 2017), inkludert frukt vekst og modning (Estrada-Johnson et al., 2017)., Derimot, når isochorismate syntase (ICS) er aktivert, chorismate kan konverteres til isochorismate og den siste kan bli forvandlet til SA. Til tross for at denne veien ble først identifisert i bakterier, det er også i dag godt etablert i planter (Wildermuth et al. I 2001; Uppalapati et al., 2007; Catinot et al., 2008; Garcion et al., 2008; Abreu og Munné-Bosch, 2009). Til slutt, tryptofan kan slå inn tryptamine av enzymet tryptofan decarboxylase (TDC)., Selv om dette er en annen av flere ruter av IAA biosyntese (tryptamine blir til indole-3-acetaldehyde og senere i IAA), TDC har også vært foreslått å være den første rate-begrense enzym i melatonin-veien (Kang et al., 2007). TDC ble først identifisert i Apocynaceae familie (De Luca et al., 1989) for å være senere beskrevet i en rekke plante-systemer (Byeon et al., 2012, 2014; Zhao et al., 2013; Wei et al., 2018)., Etter decarboxylation, tryptamine omdannes til serotonin av tryptamine-5-hydroksylase, og deretter serotonin forvandles til N-acetyl-serotonin å endelig gi melatonin i cytosol (Figur 1).
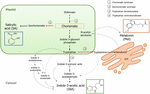
Figur 1. Biosyntese av indole-3-eddiksyre (IAA), salicylic acid (SA), og melatonin (Mel) fra chorismate. Alle tre forbindelser dele en felles endelige forløper (chorismate, kvadrerte i oransje) fra shikimate veien., Store avledning poeng er angitt med røde piler og tilsvarende enzymer er nummerert i en nedstigning divergens ordre fra shikimate. Fet stiplede pilene indikerer mer enn ett trinn mellom forbindelsene. Tynne stiplete piler indikerer trasé som fortsatt ikke er godt definert.
– Tasten-viderekobling av enzymer som fører til syntese av chorismate-veien hormoner har nylig blitt identifisert i kjøttfulle frukter (Figur 1). ICS, et uttrykk som har vist seg å være avgjørende for biotiske stress toleranse (Garcion et al.,, 2008; Zeng og Han, 2010), har blitt identifisert i tomater og epler. Zhu et al. (2016) rapporterte overexpression av ICS1 (isochorismate syntase 1) under cold storage forhold i tomat, mens Zhang et al. (2017) fant aktivering av en transkripsjonsfaktor som er involvert i patogen-relaterte SA signaliserer vei inducing ICS uttrykk i epler. Forbedret TDC aktiviteten har vært rapportert å forekomme i umoden pepper frukt ved infeksjon av patogener gjennom økt TDC1 og TDC2 uttrykk (Park et al., 2013) – og i vekst scenen i mulberry frukt (Wang et al., 2016)., Til slutt, TA har blitt identifisert i grapevine, både i pre – og véraison (Böttcher et al., 2013; Gouthu og Deluc, 2015, henholdsvis), så vel som under jordbær modning (Estrada-Johnson et al., 2017). Det er verdt å merke seg imidlertid at gener og enzymer som er beskrevet i climacteric frukt (dvs., ICS) har ikke blitt identifisert i ikke-climacteric frukt (dvs., TDC og TA) og vice versa; derfor, videre studier er viktig for å fylle disse kunnskapsmangler og bedre forstå hvordan disse avledning poeng er regulert i fellesskap i løpet av frukt modning.,
Rollen Chorismate-Avledet Phytohormoner i Climacteric og Ikke-Climacteric Frukt
Unraveling mekanismer av frukt utviklingen har vært en av de store utfordringene i de siste agronomi forskning for sin økonomiske konsekvenser. I denne sammenheng, phytohormoner har blitt pekt ut som ansvarlig drivere av frukt modning, spesielt etylen og ABA i climacteric og ikke-climacteric frukt, henholdsvis. Men at disse phytohormoner kunne regulere frukt utviklingen alene var snart vist seg å være altfor enkelt., Etter omfattende forskning og med forbedring i analytisk kjemi og molekylærbiologiske teknikker, flere andre hormoner har blitt bekreftet som potensielle regulatorer av frukt utvikling og modning, inkludert chorismate-avledet phytohormoner.
Auxins Cross-Talk Med Andre Hormoner i Løpet av Frukt Satt, Vekst og Modning
Auxins er en gruppe av plante-hormon som spiller en avgjørende rolle i frukt utvikling, både utøve sin egen innflytelse og modulerende uttrykk for andre phytohormoner., Endogene innholdet av IAA er spesielt høy på frukt satt og ved første gangs vekst utviklingsstadier, etter som IAA beløp har en tendens til å avslå før modning debut, både i climacteric (Zaharah et al., 2012) og ikke-climacteric frukt (Symons et al., 2012; Teribia et al., 2016), med tilsynelatende noen unntak, som ferskener (Tatsuki et al., 2013) og litt plomme varianter (El-Sharkawy et al., 2014; Figur 2A). Det har blitt demonstrert at IAA er involvert i frukt satt innvielse i kombinasjon med gibberellins (Mezzetti et al., 2004; Serrani et al., 2010; Bermejo et al., 2018; Hu et al.,, 2018). Nedskrivning av IAA biosyntese eller signaliserer vanligvis fører til frukt parthenocarpy, men det kan også føre til unormal modning i noen frukter (Wang et al., 2005; Liu J. et al., 2018; Reig et al., 2018). Høyt innhold av IAA i innledende faser av frukt utviklingen fremme frukt vekst på grunn av auxin underforstått i celledeling i kombinasjon med cytokinins og i kontroll av celle-utvidelsen i kombinasjon med gibberellins (Liao et al., 2018)., I løpet av denne perioden, hormonelle krysstale mellom auxins og gibberellins i tillegg tillater normal frukt forme i en finjustert mediert regulering av Auxin Svar Faktorer (ARFs; Liao et al., 2018; Liu S. et al., 2018).
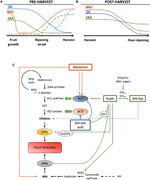
Figur 2. Rollen IAA, SA, og Mel under utviklingen av climacteric og ikke-climacteric frukt. Modellen oppsummerer vekselsvirkningene av IAA, SA, og Mel under modning av climacteric og ikke-climacteric frukt under (A) pre – og (B) post-harvest., Stiplede linjer viser alternative dynamics av phytohormone innholdet i noen frukter (se teksten for diskusjon). (C) Oversikt over samspillet av IAA, SA, og Mel med etylen og abscisic syre (ABA) biosyntese i climacteric og ikke-climacteric frukt. Auxin er en positiv regulator av etylen biosyntese av aktivering av ACC syntase gener (ACS), og inducing uttrykk for Etylen Responsive Faktorer (ERFs). Auxin også undertrykker ABA produksjon av 9-cis-epoxycarotenoid dioxygenase (NCED) undertrykkelse og hemme ABA-responsive stress gener (ARS) av ARF2 protein., Endogene IAA innholdet kan bli redusert med GH3 proteiner, syntese av som er fremmet av etylen, ABA og sukker. Usammenhengende linje som viser den mulige rollen IAA av IAA-Asp som en modning faktor styrking av etylen i noen frukter. Melatonin forbedrer ABA og etylen produksjon, selv om ACS kan bli hemmet avhengig av melatonin innholdet. SA, fungerer som en hemmer av etylen repressing ACC oksidase (ACO) gener, selv om det kan fremme uttrykk for ACS., MTA, 50-methylthioadenosine, S-AdoMet, S-adenosyl-metionin; SAM syntase, S-adenosyl-metionin syntase; ACC-1-aminocyclopropane-1-karboksylsyre; IPP, isopentenyl diphosphate.
Reduksjoner av endogene IAA innholdet har vært rapportert å forekomme før utbruddet av modning i flere frukter. Disse reduksjonene har vært knyttet til IAA konjugasjon med asparaginsyre (IAA-Asp) av IAA-amido synthetases, GH3-1 og GH3-2 (Figur 2C)., Faktisk, GH3-1 og GH3-2 viste høyere uttrykk i tidlig frukt utvikling og spesielt under modning initiation både i climacteric frukt, som for eksempel tomater (Sravankumar et al., 2018) og epler (Onik et al., 2018), så vel som i ikke-climacteric frukt, som drue bær (Böttcher et al., 2010, 2011) og bringebær (Bernales et al., 2019)., Det er interessant drue bær viste forbedret GH3-1 uttrykk etter ABA og ethephon program, som kan forklare involvering av etylen i kontroll av IAA innholdet etter utbruddet av modning, selv i ikke-climacteric frukt (Böttcher et al., 2010). Faktisk, flere studier markere tett samspill mellom auxins og etylen i frukt modning, med en gjensidig påvirkning mellom dem (Tadiello et al., 2016a; Busatto et al., 2017)., For climacteric frukt, økt innhold av IAA er nødvendig for å aktivere uttrykk for ACC syntase gener (ACS), koding ACC syntase, som, i sin tur, vil føre til produksjon av etylen som utløser den modningsprosessen (Figur 2C; Tatsuki et al., 2013). Frukt som tomater (Li et al., 2016) eller bananer (Choudhury et al.,, 2008), som vanligvis oppleve redusert IAA innholdet før utbruddet av modning, viser en modning forsinkelse når IAA eller analogs er exogenously brukt, mens frukt som fersken, som IAA innholdet øke gradvis til det er helt modne, viser akselerert modning når auxins er brukt (Tadiello et al., 2016b). Derfor, en tett, komplekse og differensial regulering av auxin-etylen vekselsvirkningene må finnes i ulike frukter i løpet av preharvest modning., Videre, IAA innholdet nedgang før modning utbruddet i noen ikke-climacteric frukt til de-undertrykke 9-cis-epoxycarotenoid dioxygenase (NCED) og start ABA syntese (Figur 2C; Jia et al., 2016). Det er interessant at denne prosessen er også formidlet av IAA bøyde gjennom forbedret GH3-1 aktivitet (Böttcher et al., 2010). Likevel, Estrada-Johnson et al. (2017) viste at selv om IAA innholdet nedgang i løpet av modne jordbær, uttrykk for FaAux/IAA og FaARF gen familier er indusert i rødt beholdere, noe som tyder på involvering av auxin signalering i fullt modne frukter.,
Under post-harvest, auxin innholdet forblir vanligvis invariant eller har en tendens til å redusere på grunn av oksidative prosesser som kan gi opphav til små, men progressiv reduksjon i den endogene innholdet av IAA (Figur 2B). Auxin behandlinger etter innhøsting forsinkelse over-modning i noen frukter (Chen et al., 2016; Mo et al., 2017) og øke innholdet av noen organiske syrer, opprettholde frukt surhet (Li et al., 2017), noe som tyder på auxin spiller også en betydelig rolle i kontroll av frukt modning under post-harvest.,
Flere Roller av salisylsyre i Løpet av Frukt Utvikling og Modning
salisylsyre er en annen chorismate-avledet phytohormone som har vært mest knyttet til dens beskyttende effekt under biotiske stress å kontrollere preharvest og post-harvest tap avledet fra patogen frukt infeksjon (Babalar et al., 2007; Cao et al., 2013). Generelt, endogen innholdet av gratis SA er høyere i begynnelsen av frukt utvikling og for deretter å avta gradvis (Oikawa et al., 2015). Lu et al. (2015) rapporterte en sekundær økning i SA under den andre vekst fase av fersken frukt., Eksogene anvendelse av SA i bananer forsinket modning prosess å redusere respiratory burst og papirmasse å skrelle forhold, samt å redusere aktiviteten av enzymer som er i slekt å cellevegg fornedrelse og antioksidant system (Srivastava og Dwivedi, 2000). Videre behandling av søte kirsebær trær med salicylic og acetylsalisylsyre (ASA) forbedret vekt, fasthet og farge av kirsebær på kommersiell fase (Giménez et al., 2014). Eksogene anvendelse av SA eller metyl salicylate (MeSA) under post-harvest, forsinket over-modning i frukter som kiwi (Zhang et al.,, 2003), og søte kirsebær (Valero et al., 2011). Videre behandlinger med salicylic eller ASA lindres chilling skader av granatepler (Sayyari et al., 2011), tomater (Aghdam et al., 2014), og avokado (Glowacz et al., 2017). SA samhandler også med etylen gjennom hemming av ACC oksidase (ACO) uttrykk (Shi og Zhang, 2012). Overraskende, økt innhold av ACS uttrykk i pærer er også funnet etter SA program i en dose-regulert måte (Shi et al., 2013; Shi og Zhang, 2014). Derfor, regulering av etylen produksjon avhengig SA er fint regulert (Figur 2C).,
Melatonin: En ny Regulator for Kontroll av Frukt-Utvikling
Melatonin er også en plante hormon fra chorismate-avledet vei som har fått mye oppmerksomhet i de senere år (Hernández-Ruiz et al., 2005; Arnao og Hernández-Ruiz, 2018; Sharif et al., 2018). Nylig, melatonin har vært påpekt som en viktig faktor for frukt sett, på en lignende måte til IAA effekter (Figur 2A; Liu S. et al., 2018). I tillegg, biologiske produksjonen av denne phytohormone har blitt observert (Kolář et al., 1997)., Under på-treet modning, druer behandlet med melatonin viste høyere synkron og økt vekt når den er fullt modner (Meng et al., 2015). Videre, en annen nylig studie av Xu et al. (2018) beskrevet at melatonin kan fremme drue berry modning gjennom sin interaksjon med ABA og etylen, samt hydrogen peroxide. Det har blitt vist at melatonin gir chilling toleranse i løpet av frukt cold storage, som synes å være mest knyttet til sin antioksidant aktivitet (Cao et al., 2018; Aghdam et al., 2019)., Under post-harvest, tomater viste økt etylen-utslipp etter påføring av melatonin på 50 µM (Sun et al., 2015), i motsetning til pærer, hvor eksogene anvendelse av melatonin på 100 µM hemmet etylen produksjon (Zhai et al., 2018), eller banan, frukt, der melatonin på 50-200 µM hemmet uttrykk for ACS og ACO (Figur 2C; Hu et al., 2017). Selv om ulike virkninger av melatonin kan programmet være knyttet til en dose-effekt, for videre forskning er nødvendig for å bedre forstå betydningen av melatonin i kontroll av frukt utvikling.,
Cross-Talk av Chorismate-Avledet Phytohormoner i Kontroll av Frukt-Utvikling
Hormonelle krysstale mellom chorismate-avledet phytohormoner har vist seg å oppstå i løpet av frukt utvikling og modning. Breitel et al. (2016) fant at auxins kan samhandle med SA gjennom ARF2 (Figur 2C), og at overexpression av ARF2 i tomat resulterte i lavere innhold av SA og en betydelig modning forsinkelse., Dette indikerer at auxins kan være å begrense SA produksjon i løpet av frukt utvikling, noe som kan forklare en trade-off mellom frukt vekst (mediert av auxin) og aktivering av biotiske forsvar (mediert av SA). Videre krever anvendelse av SA på papaya frukt resultert i en endret uttrykk av flere IAA gener, noen av dem blir nedregulert mens andre opp-regulert (Liu et al., 2017)., I dette tilfellet, resultatene var usikker på hva som egentlig skjer endogenously i den antatte cross-talk mellom auxin og SA i løpet av frukt modning og videre forskning er nødvendig i dette og andre climacteric frukt. I alle fall er det klart at IAA og SA er nært knyttet til hverandre, ikke bare metabolically, men også funksjonelt. Til slutt, flere studier har rapportert påvirkning av melatonin på IAA og SA biosyntese og/eller signalering (anmeldt i Arnao og Hernández-Ruiz, 2018), selv om vår kunnskap, ingen av disse studiene ble utført verken i climacteric heller i ikke-climacteric frukt.,
Modulering av Bioaktive Forbindelser Biosyntese av Chorismate-Avledet Phytohormoner
Bioaktive forbindelser (inkludert fenoliske forbindelser, isoprenoids, og vitaminene) har ikke bare vært mye undersøkt som ansvarlige for bestemte organoleptiske egenskaper av matvarer, men også for sine beskyttende effekter på humane celler mot oksidativt prosesser i utviklingen av nevrodegenerativ og hjerte-og karsykdommer og visse typer kreft (Liu et al. I 1999; Mueller et al., 2010; Stør og Ronnenberg, 2010; Björkman et al., 2011)., Fenoliske forbindelser som i betydelig grad bidrar til å formidle spesifikke smaker, som for eksempel tanniner – ansvarlig for bitterhet eller astringency smak av visse frukter, og farger, slik som anthocyanin pigmenter – ansvarlig for rødt, blått og lilla frukt farger (Croteau et al., 2000). Relevansen av fenoliske forbindelser til konsum har vært forbundet med en beskyttende effekt mot oksidative prosesser i forhold til hjerte-og sentral-nervesystemet helse, samt en redusert risiko for kreft i mage-tarmkanalen (Björkman et al., 2011)., Karotenoider, tetraterpenes som tilhører isoprenoids, spille en rolle i beskyttelse mot foto-oksidative prosesser og som organiske pigmenter, de er ansvarlig for den oransje-gul farge av frukt (Tapiero et al., 2004). Kosttilskudd karotenoider er tenkt å gi helsemessige fordeler i å redusere risikoen for øye sykdom og visse typer kreft på grunn av deres rolle som antioksidanter (Johnson, 2002). Vitamin C og E, inkludert ascorbic acid og tokoferoler, henholdsvis, er viktige ernæringsmessige kvalitet faktorer i frukt med mange biologiske aktiviteter i mennesker (Miret og Müller, 2017)., Tatt i betraktning de relevante rollen som alle disse bioaktive forbindelser som helsefremmende stoffer i frukt, modulasjonen av deres akkumulering er av avgjørende betydning i begge climacteric og ikke-climacteric frukt (Vasconsuelo og Boland, 2007). De fleste av disse bioaktive forbindelser er samlet på et høyt nivå i løpet av frukt modning, når mindre velsmakende grønne frukter snakke inn i en ernæringsmessig rik, farget, og velsmakende frukt (Daood et al., 1996; Ranalli et al., 1998; Dumas et al., 2003; Singh et al., 2011)., Men, når startet, modning kan ikke bli stoppet og generelt fører til over-modning som negativt påvirker frukt kvalitet (Kumar et al., 2014). Derfor, minimere post-harvest frukt ødeleggelse mens å skaffe høyt innhold av bioaktive forbindelser er fortsatt en av de største utfordringer å løse.
Auxin har blitt identifisert som en viktig regulator av karotenoid biosyntese under modning av climacteric tomat frukt. Modning i tomat er knyttet til nedbrytning av klorofyll og skift av xanthophylls å carotenes (β-karoten og lycopene) (Fraser et al., 1994)., IAA vises til forsinkelse tomat modning av repressing etylen og flere oppstrøms karotenoid transkripsjoner inkludert Psy, Ziso, Pds, Critiso samt Chlases 1-3 og, på den annen side, å fremme akkumulering av β-Lyc1 og Crtr-β1 transkripsjoner, noe som resulterer i høyere innhold av xanthophylls og klorofyll a (Su et al., 2015). I tillegg, i noen ikke-climacteric frukt som kirsebær og drue bær auxin virker også til å modulere anthocyanin biosyntese og dermed kontroll frukt modning prosesser. Teribia et al., (2016) rapporterte en negativ korrelasjon mellom IAA og anthocyanin innholdet indikerer at anthocyanin akkumulering starter når IAA innholdet nedgang. Videre vil anvendelse av syntetisk auxin-som sammensatte benzothiazole-2-oxyacetic syre (BTOA) forsinket opp-regulering av gener som fremmer enzymer av anthocyanin biosyntese som chalcone syntase og UDP-glukose:flavonoid 3-O-glucosyltransferase i druen bær (Davies et al., 1997).,
Pre – og post-harvest-behandlinger med salisylater, inkludert SA og dets derivater ASA og MeSA, har blitt rapportert til å regulere bioaktive forbindelser akkumulering fører til forbedret antioksidant aktivitet i både climacteric og ikke-climacteric frukt. Flere frukt som plomme, kirsebær og aprikoser er kjent for å ha et kort liv med en rask forverring i kvalitet etter innhøsting. Derfor er det en konstant jakt etter behandlinger som forbedre og vedlikeholde frukt kvalitet, og spesielt innhold av bioaktive forbindelser med gunstige helseeffekter., Behandlinger av plommer med SA, ASA, og MeSA resulterte i signifikant høyere innholdet av askorbinsyre, anthocyanins og fenoliske forbindelser både på høst og etter langvarig kulde lagring (40 dager ved 4°C; Martínez-Esplá et al., 2017). Lignende resultater kan observeres for preharvest behandlinger med SA, ASA, og MeSA i søte kirsebær (Giménez et al., 2014, 2015, 2017). Videre, post-harvest behandling med SA, ASA, eller MeSA vedlikeholdt totalt fenoliske innhold samt anthocyanin innholdet i kaldt lagring i granateple (Sayyari et al., 2011), søte kirsebær (Valero et al.,, 2011), cornelian cherry (Dokhanieh et al., 2013), og aprikos (Wang et al., 2015). Disse resultatene tyder på at salisylater kan være involvert i aktivering av fenylalanin ammoniakk lyase, som er den viktigste enzymet er involvert i biosynthesic vei av fenoliske forbindelser (Martinez-Esplá et al., 2017).
Nylig, melatonin har vist seg å regulere frukt modning og modulere bioaktive forbindelser. Post-harvest behandling med melatonin i fersken frukt økt chilling toleranse ved å aktivere antioksidant-systemer., Chilling skaden oppstår i fersken frukt under lav temperatur lagring (mellom 2 og 7.6°C) preget av kjøtt bruning, unormal modning og høyere følsomhet til forfall (Lurie og Crisosto, 2005). Cao et al. (2018) rapporterte at peaches behandlet med melatonin viste høyere transkripsjonen overflod av ascorbic acid biosyntese gener PpGME, PpGPP, og PpGLDH til syvende og PpGMPH på 21st og 28. dag lagringsplass i forhold til å styre noe som resulterer i økt ascorbic acid innhold., Videre, melatonin indusert økning i aktiviteter av G6PDH, SKDH og PAL, som er viktige enzymer for fenoliske forbindelser biosyntese. Forfatterne foreslår at melatonin behandling beskytter fersken frukt til en viss grad fra chilling skade ved spesielt å aktivere biosyntese av fenoliske forbindelser (Gao et al., 2018). I tillegg, en label-fri differensial proteomics analyse avslørte effekten av melatonin på å fremme frukt modning og anthocyanin akkumulering ved post-harvest i tomat frukt (Sun et al., 2016)., Forfatterne rapporterte at eksogene melatonin økt åtte enzymer knyttet til anthocyanin vei, herunder blant annet flavonol-3-hydroksylase, flavanone 3 beta-hydroksylase, anthocyanidin syntase/leucoanthocyanidin dioxygenase, og anthocyanidin 3-O-glucosyltransferase. Videre, post-harvest behandling med melatonin økt total fenol og anthocyanins i jordbær frukt (Aghdam og Fard, 2016), og forsinket tap av total fenol, flavonoider og anthocyanins i litchi frukt (Zhang et al., 2018)., Behandling med melatonin økte innholdet av fenoler, anthocyanins og flavonoider i druen bær (Xu et al., 2017). I tillegg, melatonin har vært rapportert å øke lycopene oppsamling og produksjon av etylen i tomater, noe som tyder på at melatonin kan øke innholdet av lycopene ved å påvirke etylen biosyntese og signalering (Sun et al., 2015)., Deltakelse av etylen i melatonin-indusert regulering av bioaktive forbindelser har også blitt observert i druen bær, der double block behandling av etylen viste redusert effekten av melatonin på polyphenol innhold (Xu et al., 2017).
Konklusjon
Det er konkludert med at chorismate-avledet phytohormoner, inkludert auxin, SA og melatonin, ikke bare dele en felles forløper, men spille viktige roller i regulering av frukt vekst og modning., En metabolsk og funksjonelle cross-talk mellom dem og med andre phytohormoner oppstår i en spatiotemporal måte å fint regulere utviklingen av climacteric og ikke-climacteric frukt. Forskjellene i dynamikken innen climacteric og ikke-climacteric frukt bevis for at responsen chorismate-avledet hormoner er ikke universell, men heller sterkt arts-spesifikke. Videre, chorismate-avledet phytohormoner også modulere akkumulering av bioaktive forbindelser, og dermed påvirke frukt kvalitet., Mer forskning er imidlertid nødvendig for å bedre forstå hvordan du (i) andre hormoner, som for eksempel etylen og ABA, modulere deres biosyntese å studere viktige metabolske avledning poeng, (ii) de samhandler på funksjonelle og molekylært nivå, og (iii) de i fellesskap modulere bioaktive forbindelser biosyntese og dermed innflytelse, ikke bare frukt vekst og modning, men også kvaliteten på climacteric og ikke-climacteric frukt.
Forfatter Bidrag
SM-B unnfanget og designet gjennomgang med hjelp av MP-L. MP-L, PM, MM, og SM-B skrev manus., MP-L forberedt tall ved hjelp av PM. Alle forfatterne har bidratt til diskusjon av ideer, revidert og godkjent det siste manuskriptet.
Midler
Dette arbeidet ble finansiert av en ICREA Akademia utmerkelse som gis til SM-B ved den katalanske Regjeringen.
interessekonflikt Uttalelse
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.
Böttcher, C., Boss, P. K., og Davies, C. (2011)., Acyl substrat preferanser av en IAA-amido synthetase konto for variasjoner i druen (Vitis vinifera L.) berry modning forårsaket av ulike auxinic forbindelser som indikerer viktigheten av auxin konjugasjon i anlegget utvikling. J. Exp. Robot. 62, 4267-4280. doi: 10.1093/jxb/err134
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., og Sivakumar, D. (2017)., Vedlikeholde postharvest kvaliteten på kalde lagret ‘Hass’ avokado ved å endre fettsyrer innhold og sammensetning med bruk av naturlige flyktige forbindelser – methyl jasmonate og metylsalisylat. J. Sci. Mat. Agric. 97, 5186-5193. doi: 10.1002/jsfa.8400
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Zeng, W., og Han, S. Y. (2010). En fremtredende rolle flagellin reseptor FLAGELLIN-SENSING2 i formidling stomatal svar på Pseudomonas syringae pv tomat DC3000 i Arabidopsis. Anlegget Physiol. 153, 1188-1198. doi: 10.1104/s. 110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















