Biologie pentru Majors I
Învățării
- să Descrie rolul ARN polimeraza
Această secțiune se va extinde la rolul specific al ARN-polimeraze timpul de transcriere. Citiți mai departe pentru a afla rolul ARN polimerazelor în fiecare etapă a transcrierii.,spre deosebire de polimeraza procariotă care se poate lega singură de un șablon ADN, eucariotele necesită mai multe alte proteine, numite factori de transcripție, pentru a se lega mai întâi de regiunea promotorului și apoi pentru a ajuta la recrutarea polimerazei adecvate.
cele trei polimeraze ARN eucariote
caracteristicile sintezei ARNm eucariote sunt semnificativ mai complexe decât cele ale procariotelor. În loc de o singură polimerază care cuprinde cinci subunități, eucariotele au trei polimeraze care sunt fiecare alcătuite din 10 subunități sau mai multe., Fiecare polimerază eucariotă necesită, de asemenea, un set distinct de factori de transcripție pentru ao aduce la șablonul ADN.
ARN polimeraza I este localizată în nucleol, o substructură nucleară specializată în care ARN ribozomal (ARNr) este transcris, prelucrat și asamblat în ribozomi (Tabelul 1). Moleculele ARNr sunt considerate ARNr structurale deoarece au un rol celular, dar nu sunt traduse în proteine. ARNr sunt componente ale ribozomului și sunt esențiale pentru procesul de traducere. ARN polimeraza i sintetizează toate ARNr-urile, cu excepția moleculei rRNA 5s., Denumirea ” S „se aplică unităților” Svedberg”, o valoare nonadditivă care caracterizează viteza cu care o particulă sedimentează în timpul centrifugării.
| Tabelul 1.,f Transcription | α-Amanitin Sensitivity | |||
|---|---|---|---|---|
| I | Nucleolus | All rRNAs except 5S rRNA | Insensitive | |
| II | Nucleus | All protein-coding nuclear pre-mRNAs | Extremely sensitive | |
| III | Nucleus | 5S rRNA, tRNAs, and small nuclear RNAs | Moderately sensitive | |
RNA polymerase II is located in the nucleus and synthesizes all protein-coding nuclear pre-mRNAs., Pre-ARNm eucariote sunt supuse unei prelucrări extinse după transcriere, dar înainte de traducere (Figura 1). Pentru claritate, discuția acestui modul despre transcriere și traducere în eucariote va folosi termenul „mRNAs” pentru a descrie numai moleculele mature, prelucrate, care sunt gata de a fi traduse. ARN polimeraza II este responsabilă pentru transcrierea majorității covârșitoare a genelor eucariote.

Figura 1. ARNm eucariot conține introni care trebuie îmbinați. De asemenea, se adaugă un capac de 5′ și 3′ coadă poli-A.,
ARN polimeraza III este, de asemenea, localizată în nucleu. Această polimerază transcrie o varietate de ARN-uri structurale care includ 5S pre-rRNA, pre-ARN-uri de transfer (pre-tRNAs) și pre-ARN-uri nucleare mici. TRNAs au un rol critic în traducere; ele servesc ca molecule de adaptare între șablonul ARNm și lanțul polipeptidic în creștere. ARN-urile nucleare mici au o varietate de funcții, inclusiv „splicing” pre-mRNAs și reglarea factorilor de transcripție.,
Un om de știință care caracterizează o nouă genă poate determina care polimerazei transcrie prin testarea dacă gena este exprimat în prezența unei anumite ciuperci otravă, α-amanitin (Tabelul 1). Interesant este că α-amanitina produsă de Amanita phalloides, ciuperca de moarte, afectează foarte diferit cele trei polimeraze. ARN polimeraza i este complet insensibilă la α-amanitină, ceea ce înseamnă că polimeraza poate transcrie ADN-ul in vitro în prezența acestei otrăviri. În schimb, ARN polimeraza II este extrem de sensibilă la α-amanitină, iar ARN polimeraza III este moderat sensibilă., Cunoașterea polimerazei transcriptoare poate determina un cercetător în funcția generală a genei studiate. Deoarece ARN polimeraza II transcrie marea majoritate a genelor, ne vom concentra pe această polimerază în discuțiile noastre ulterioare despre factorii și promotorii transcripției eucariote.promotorii ARN polimerazei II și factorii de transcripție promotorii Eucarioți sunt mult mai mari și mai complicați decât promotorii procarioți. Cu toate acestea, ambele au o secvență similară cu secvența -10 de procariote., În eucariote, această secvență se numește caseta TATA și are secvența de consens TATAAA pe componenta de codificare. Acesta este situat la -25 la -35 baze în raport cu site-ul de inițiere (+1) (Figura 2). Această secvență nu este identică cu cutia E. coli -10, dar conservă elementul bogat A–T. Termostabilitatea legăturilor A-T este scăzută și acest lucru ajută șablonul ADN să se relaxeze local în pregătirea transcrierii.,
în loc de factorul σ simplu care ajută la legarea ARN polimerazei procariote de promotorul său, eucariotele asamblează un complex de factori de transcripție necesari pentru a recruta ARN polimeraza II la o genă care codifică proteina. Factorii de transcripție care se leagă de promotor se numesc factori de transcripție bazală. Acești factori bazali sunt toți numiți TFII (pentru factorul de transcripție/polimeraza II) plus o literă suplimentară (A-J). Complexul de bază este TFIID, care include o proteină care leagă TATA (TBP)., Alți factori de transcripție sistematic cădea în loc pe ADN-ul șablon, fiecare cu o continuare stabilizarea pre-inițiere complexe și contribuie la recrutarea de ARN polimeraza II.
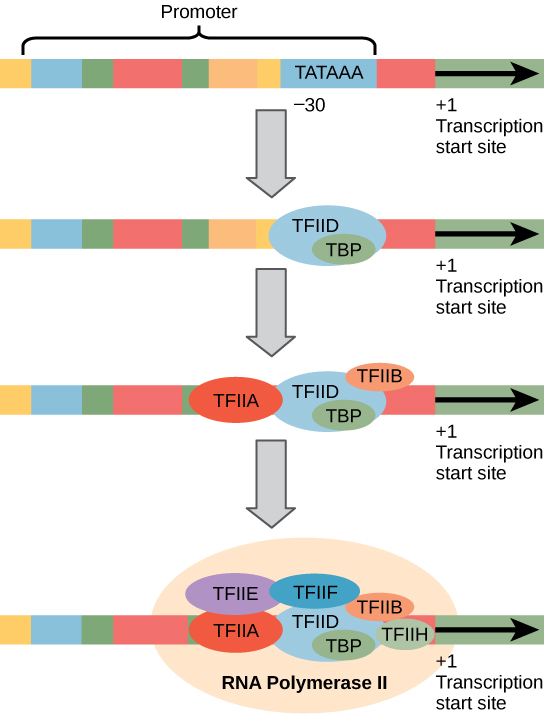
Figura 2. Este prezentat un promotor generalizat al unei gene transcrise de ARN polimeraza II. Factorii de transcriere recunosc promotorul. ARN polimeraza II se leagă apoi și formează complexul de inițiere a transcripției.,
întrebare practică
un om de știință îmbină un promotor eucariot în fața unei gene bacteriene și introduce gena într-un cromozom bacterian. Te-ai aștepta ca bacteriile să transcrie gena?unii promotori eucarioți au, de asemenea, o cutie CAAT conservată (GGCCAATCT) la aproximativ -80. Mai în amonte de TATA box, eucariote promotori, de asemenea, poate conține unul sau mai multe GC-bogat cutii (GGCG) sau octamer cutii (ATTTGCAT)., Aceste elemente leagă factorii celulari care măresc eficiența inițierii transcripției și sunt adesea identificați în gene mai „active” care sunt în mod constant exprimate de celulă.factorii de transcripție bazali sunt cruciali în formarea unui complex de preinițiere pe șablonul ADN care recrutează ulterior ARN polimeraza II pentru inițierea transcripției. Complexitatea transcrierii eucariote nu se termină cu polimerazele și promotorii., O armată de alți factori de transcripție, care se leagă de amplificatorii și amortizoarele din amonte, ajută, de asemenea, la reglarea frecvenței cu care pre-ARNm este sintetizat dintr-o genă. Amplificatoarele și amortizoarele de zgomot afectează eficiența transcrierii, dar nu sunt necesare pentru ca transcrierea să continue.
evoluția promotorilor
evoluția genelor poate fi un concept familiar. Mutațiile pot apărea în gene în timpul replicării ADN-ului, iar rezultatul poate sau nu poate fi benefic pentru celulă., Prin modificarea unei enzime, proteine structurale, sau un alt factor, procesul de mutație poate transforma funcții sau caracteristici fizice. Cu toate acestea, promotorii eucarioți și alte secvențe de reglementare a genelor pot evolua, de asemenea. De exemplu, luați în considerare o genă care, de-a lungul multor generații, devine mai valoroasă pentru celulă. Poate că gena codifică o proteină structurală pe care celula trebuie să o sintetizeze din abundență pentru o anumită funcție. În acest caz, ar fi benefic pentru celulă ca promotorul genei să recruteze mai eficient factorii de transcripție și să crească expresia genelor.,
oamenii de știință care examinează evoluția secvențelor promotorului au raportat rezultate diferite. În parte, acest lucru se datorează faptului că este dificil să se deducă exact unde începe și se termină un promotor eucariot. Unii promotori apar în cadrul genelor; alții sunt localizați foarte departe în amonte sau chiar în aval de genele pe care le reglează. Cu toate acestea, atunci când cercetătorii au limitat examinarea lor umane de bază promotor secvențe care au fost definite experimental ca secvențe care leagă preinitiation complexe, au descoperit că promotorii evolua chiar mai repede decât de proteine-codare gene.,
este încă neclar modul în care evoluția promotorului ar putea corespunde evoluției oamenilor sau a altor organisme superioare. Cu toate acestea, evoluția unui promotor pentru a face în mod eficient mai mult sau mai puțin dintr-un produs genic dat este o alternativă interesantă la evoluția genelor în sine.
structuri promotoare pentru polimerazele ARN I și III
procesele de aducere a polimerazelor ARN I și III la șablonul ADN implică colecții puțin mai complexe de factori de transcripție, dar tema generală este aceeași.,elementele promotoare conservate pentru genele transcrise de polimerazele I și III diferă de cele transcrise de ARN polimeraza II. ARN polimeraza I transcrie gene care au două secvențe promotoare bogate în GC în regiunea -45 până la +20. Numai aceste secvențe sunt suficiente pentru inițierea transcripției, dar promotorii cu secvențe suplimentare în regiunea de la -180 la -105 în amonte de locul inițierii vor spori și mai mult inițierea. Genele care sunt transcrise de ARN polimeraza III au promotori sau promotori în amonte care apar în genele în sine.,transcrierea eucariotă este un proces bine reglementat care necesită o varietate de proteine pentru a interacționa între ele și cu catena ADN. Deși procesul de transcriere în eucariote implică o investiție metabolică mai mare decât în procariote, se asigură că celula transcrie exact pre-ARN-urile de care are nevoie pentru sinteza proteinelor.,în urma formării complexului de preinițiere, polimeraza este eliberată de ceilalți factori de transcripție, iar alungirea este permisă să se desfășoare așa cum se întâmplă în procariote cu polimeraza care sintetizează pre-ARNm în direcția 5′ până la 3′. După cum sa discutat anterior, ARN polimeraza II transcrie ponderea majoră a genelor eucariote, astfel încât această secțiune se va concentra pe modul în care această polimerază realizează alungirea și terminarea.,deși procesul enzimatic de alungire este în esență același în eucariote și procariote, șablonul ADN este mai complex. Când celulele eucariote nu se divid, genele lor există ca o masă difuză de ADN și proteine numite cromatină. ADN-ul este bine ambalat în jurul proteinelor histone încărcate la intervale repetate. Aceste complexe ADN-histone, denumite colectiv nucleozomi, sunt distanțate în mod regulat și includ nucleotide 146 de ADN înfășurate în jurul a opt histone ca firul în jurul unei bobine.,
Pentru polinucleotide sinteza să apară, transcrierea utilaje trebuie să se miște histones din drum de fiecare dată când se întâlnește un nucleosome. Acest lucru este realizat printr-un complex special de proteine numit FACT, care înseamnă „facilitează transcrierea cromatinei.”Acest complex trage histone departe de șablonul ADN pe măsură ce polimeraza se mișcă de-a lungul acestuia. Odată ce pre-ARNm este sintetizat, complexul fapt înlocuiește histone pentru a recrea nucleozomii.
terminarea transcrierii este diferită pentru diferitele polimeraze., Spre deosebire de procariote, alungirea prin ARN polimeraza II în eucariote are loc 1,000–2,000 nucleotide dincolo de sfârșitul genei transcrise. Această coadă pre-ARNm este ulterior îndepărtată prin scindare în timpul procesării ARNm. Pe de altă parte, ARN polimerazele I și III necesită semnale de terminare. Genele transcrise de ARN polimeraza i conțin o secvență specifică de 18 nucleotide care este recunoscută de o proteină terminală. Procesul de terminare în ARN polimeraza III implică un ac de păr mRNA similar cu terminarea rho-independentă a transcripției în procariote.,
încercați să-l
contribuie!
îmbunătățiți această paginăaflați mai mult
- H Liang și colab., „Evoluția rapidă a promotorilor de bază în genomul primatelor,” Biologie Moleculară și Evoluție 25 (2008): 1239-44. ↵















