Frontiere în Plant Science
Introducere
Corect progresie de coacere a fructelor este esențială pentru a realiza atât optimă de fructe de calitate și termen de valabilitate lung, trăsături importante care determina prețul piețelor și a profitului final. Cu toate acestea, maturarea este un proces complex în care sunt implicați mulți factori, inclusiv controlul hormonal, care reglează modificările biochimice și fiziologice care dau proprietăți organoleptice și nutriționale finale ale fructelor., În mod tradițional, fructele cărnoase au fost clasificate în funcție de procesul de maturare în fructe climacterice și non-climacterice. Fructele climacterice, cum ar fi rosii sau banane necesită o creștere în rata respirației și a producției de etilenă pentru a dezlănțui procesul de maturare într-o autocatalitice răspuns (Barry și Giovannoni, 2007; Liu et al., 2015)., Antagonistically, non-climaterice fructe, cum ar fi citrice sau struguri nu prezintă o spargere nici în etilenă, nici respirația înainte de maturare debut, și în aceste fructe, de maturare este controlată în principal de acumularea progresivă de fitohormoni acid abscisic (ABA; Rodrigo et al., 2006; Castellarin și colab., 2011). Cu toate acestea, noi studii consolidează ideea că procesul de maturare nu este guvernat doar de producerea unei fitohormoni, ci mai degrabă pare a fi rezultatul unui echilibru hormonal controlat (Symons et al., 2012; Teribia și colab., 2016; Li și colab., 2019).,auxinele, acidul salicilic (SA) și melatonina sunt fitohormoni implicați în semnalizarea și reglarea multor procese cruciale în plante. Auxinele au fost descrise pe scară largă ca regulatori de creștere și dezvoltare cu funcții multiple în plante (vezi Taylor-Teeples et al., 2016); SA declanșează răspunsul de apărare împotriva o ciupercǎ și hemi-o ciupercǎ patogeni (Loake și Grant, 2007), precum și ca având un rol important sub stres abiotic (Dong et al., 2014; Wani și colab., 2017), înflorire și controlul ciclului celular (Carswell et al., 1989; Eberhard și colab.,, 1989, respectiv); și melatonina nu numai că a fost găsită ca având funcții asemănătoare auxinei (Chen et al., 2009; Zuo și colab., 2014; Wen și colab., 2016), dar, de asemenea, sa sugerat să acționeze ca un potențial antioxidant în unele plante (Arnao și Hernández-Ruiz, 2015) și un regulator al răspunsurilor plantelor la agenții patogeni (Chen et al., 2018). Interesant, aceste trei hormoni în comun un precursor – chorismate -, astfel metabolice cross-talk apare între ele, și un număr de gene trebuie să fie fin reglată pentru a distrage chorismate metabolismul față de acești compuși., În ultimii ani, mai multe recenzii au abordat biosinteza și rolul auxinei (Paul et al., 2012), SA (Asghari și Aghdam, 2010) și melatonină (Feng și colab., 2014; Arnao și Hernández-Ruiz, 2018) în fructe. Cu toate acestea, biosinteza și funcțiile lor au fost descrise în mare parte separat. Aici vom discuta aspectele comune și diferențiate ale biosintezei, metabolismului și funcției auxinei, SA și melatoninei în creșterea și maturarea fructelor climacterice și non-climacterice., Se va pune accentul pe diversiunea metabolică a punctelor cheie în biosinteza lor din corismat, rolul de reglementare în timpul maturării fructelor climacterice și non-climacterice și rolul lor în modularea biosintezei compușilor bioactivi, care determină în mare măsură calitatea fructelor.
Chorismate Derivate Fitohormoni
Chorismate este produsul final al shikimate cale și joacă un rol-cheie în biosinteza de fitohormoni (Figura 1)., Chorismate dă naștere la aminoacizii aromatici, inclusiv triptofan, și prin mai multe reacții, inclusiv conversia triptofanului la indol-3-acid piruvic de triptofan aminotransferazei (TA); se poate genera auxinei indol-3-acetic (IAA). TA a fost propusă ca o enzimă cheie universală pentru biosinteza IAA nu numai în organele vegetative (Enríquez-Valencia et al., 2018), dar și în dezvoltarea organelor de reproducere (Reyes-Olalde et al., 2017), inclusiv creșterea și maturarea fructelor (Estrada-Johnson et al., 2017)., Prin contrast, atunci când isochorismate sintetazei (ICS) este activat, chorismate poate fi transformat într-isochorismate și aceasta din urmă poate fi transformat într-SA. În ciuda faptului că această cale a fost identificată pentru prima dată în bacterii, în prezent este bine stabilită și în plante (Wildermuth et al., 2001; Uppalapati și colab., 2007; Catinot și colab., 2008; Garcion și colab., 2008; Abreu și Munné-Bosch, 2009). În cele din urmă, triptofanul se poate transforma în triptamină prin enzima triptofan decarboxilază (TDC)., Deși aceasta este o altă cale multiplă de biosinteză IAA (triptamina se transformă în indol-3-acetaldehidă și mai târziu în IAA), TDC a fost, de asemenea, propusă a fi prima enzimă care limitează rata în calea melatoninei (Kang et al., 2007). TDC a fost identificat pentru prima dată în familia Apocynaceae (De Luca et al., 1989) pentru a fi descris mai târziu într-o serie de sisteme de plante (Byeon et al., 2012, 2014; Zhao și colab., 2013; Wei și colab., 2018)., După decarboxilare, triptamina este transformată în serotonină de către triptamină-5-hidroxilază, iar apoi serotonina se transformă în N-acetil-serotonină pentru a produce melatonină în citosol (Figura 1).
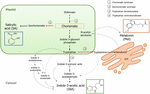
Figura 1. Biosinteza acidului indol-3-acetic (IAA), a acidului salicilic (SA) și a melatoninei (Mel) din corismat. Toți cei trei compuși au un precursor final comun (chorismate, pătrat în portocaliu) din calea shikimate., Punctele majore de deviere sunt indicate cu săgeți roșii, iar enzimele corespunzătoare sunt numerotate într-o ordine de divergență de coborâre de la shikimate. Săgețile punctate cu caractere aldine indică mai mult de un pas între compuși. Săgețile punctate subțiri indică căi care încă nu sunt bine definite.enzimele de deviere a cheilor care conduc la sinteza hormonilor căii corismate au fost identificate recent în fructele cărnoase (Figura 1). ICS, a căror expresie sa dovedit a fi crucială pentru toleranța la stres biotic (Garcion și colab.,, 2008; Zeng și He, 2010), a fost identificat în tomate și mere. Zhu și colab. (2016) raportat supraexpresia ICS1 (isochorismate sintetazei 1) în depozitare la rece în condiții de roșii, în timp ce Zhang et al. (2017) a constatat activarea unui factor de transcripție implicat în calea de semnalizare sa legată de agentul patogen care induce expresia ICS în mere. Activitatea îmbunătățită a TDC a fost raportată să apară în fructele de piper necoapte la infecția cu agenți patogeni – prin creșterea expresiei TDC1 și TDC2 (Park et al., 2013) – și în stadiul de creștere a fructelor de dud (Wang et al., 2016)., În cele din urmă, TA a fost identificat în viță de vie, atât la pre – și véraison (Böttcher et al., 2013; Gouthu și Deluc, respectiv 2015), precum și în timpul maturării căpșunilor (Estrada-Johnson et al., 2017). Este de remarcat, totuși, că genele și enzimele descrise în fructele climacterice (adică ICS) nu au fost identificate în fructele non-climacterice (adică TDC și TA) și invers; prin urmare, studiile suplimentare sunt imperative pentru a umple aceste lacune de cunoștințe și pentru a înțelege mai bine modul în care aceste puncte de deviere sunt reglementate în comun în timpul maturării fructelor.,
Rol de Chorismate Derivate Fitohormoni în Climaterice și Non-Climaterice Fructe
Deslușesc mecanismele de fructe dezvoltare a fost una dintre provocările majore în ultimii agronomie cercetări pentru implicațiile sale economice. În acest context, fitohormonii au fost evidențiați ca factori responsabili de maturare a fructelor, în special etilenă și ABA în fructele climacterice și, respectiv, non-climacterice. Cu toate acestea, că aceste fitohormoni ar putea reglementa dezvoltarea fructelor singur sa dovedit curând a fi mult prea simplu., După cercetări ample și cu îmbunătățirea chimiei analitice și a tehnicilor moleculare, mai mulți alți hormoni au fost confirmați ca potențiali regulatori ai dezvoltării și maturării fructelor, inclusiv fitohormoni derivați de corismat.auxinele sunt un grup de hormoni vegetali care joacă un rol esențial în dezvoltarea fructelor, atât exercitând propria lor influență, cât și modulând expresia altor fitohormoni., Conținutul endogen al IAA este deosebit de ridicat la stabilirea fructelor și în stadiile inițiale de dezvoltare a creșterii, după care cantitățile IAA tind să scadă înainte de debutul maturării, atât în climacteric (Zaharah et al., 2012) și fructe non-climacterice (Symons et al., 2012; Teribia și colab., 2016), cu aparent unele excepții, cum ar fi piersicile (Tatsuki et al., 2013) și unele soiuri de prune (El-Sharkawy et al., 2014; figura 2A). S-a demonstrat că IAA este implicată în inițierea setului de fructe în combinație cu gibberelinele (Mezzetti et al., 2004; Serrani și colab., 2010; Bermejo și colab., 2018; Hu și colab.,, 2018). Afectarea biosintezei sau semnalizării IAA duce, în general, la parthenocarpia fructelor, deși poate duce și la maturarea anormală a unor fructe (Wang et al., 2005; Liu J. și colab., 2018; Reig și colab., 2018). Conținutul ridicat de IAA în stadiile inițiale de dezvoltare de fructe promova creșterea fructelor datorită auxin implicarea în diviziunea celulară, în combinație cu citochinine și în controlul de expansiune de celule în combinație cu gibereline (Liao et al., 2018)., În această perioadă, crosstalk hormonal între auxine și gibberellins permite în plus modelarea normală a fructelor într-o reglare fină mediată de factorii de răspuns Auxin (ARFs; Liao et al., 2018; Liu S. și colab., 2018).
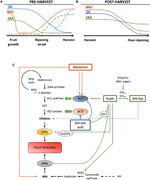
Figura 2. Rolul IAA, SA și Mel în timpul dezvoltării fructelor climacterice și non-climacterice. Modelul care rezumă interacțiunile IAA, SA și Mel în timpul maturării fructelor climacterice și non-climacterice în timpul (a) pre – și (B) post-recoltare., Liniile punctate indică dinamica alternativă a conținutului de fitohormoni în unele fructe (vezi textul pentru discuții). (C) Prezentare generală a interacțiunii IAA, SA și Mel cu biosinteza etilenei și a acidului abscisic (ABA) în fructele climacterice și non-climacterice. Auxina este un regulator pozitiv al biosintezei etilenei prin activarea genelor ACC sintază (ACS) și inducerea expresiei factorilor de răspuns la etilenă (ERFs). Auxina, de asemenea, reprimă ABA producție de 9-cis-epoxycarotenoid dioxygenase (I) represiune și inhibarea ABA-receptiv stres gene (ARS) de ARF2 proteine., Conținutul IAA endogen poate fi redus prin proteine GH3, a căror sinteză este promovată de etilenă, ABA și zaharuri. Linia discontinuă indică rolul posibil al IAA de către IAA-Asp ca factor de maturare care îmbunătățește etilena în unele fructe. Melatonina îmbunătățește producția de ABA și etilenă, deși ACS poate fi inhibată în funcție de conținutul de melatonină. Sa acționează ca un inhibitor al etilenei care reprimă genele ACC oxidază (ACO), deși poate promova expresia ACS., MTA, 50-methylthioadenosine; S-AdoMet, S-adenosyl-metionină; SAM-sintetazei, S-adenosyl-metionină-sintetazei; ACC, 1-aminocyclopropane-1-carboxilic; IPP, isopentenyl difosfat.au fost raportate reduceri ale conținutului endogen de IAA înainte de debutul maturării la mai multe fructe. Aceste reduceri au fost legate de conjugarea IAA cu acid aspartic (IAA-Asp) de către IAA-amido sintetaze, GH3-1 și GH3-2 (Figura 2c)., Într-adevăr, GH3-1 și GH3-2 au arătat o expresie mai mare în timpul dezvoltării timpurii a fructelor și mai ales în timpul inițierii maturării atât în fructele climacterice, cum ar fi roșiile (Sravankumar et al., 2018) și mere (Onik et al., 2018), precum și în fructele non-climacterice, cum ar fi boabele de struguri (Böttcher et al., 2010, 2011) și zmeură (Bernales et al., 2019)., Interesant este că boabele de struguri au prezentat o expresie îmbunătățită a GH3-1 după aplicarea ABA și etefon, ceea ce ar putea explica implicarea etilenei în controlul conținutului IAA după debutul maturării, chiar și în fructele non-climacterice (Böttcher et al., 2010). De fapt, mai multe studii evidențiază interacțiunea strânsă dintre auxine și etilenă în maturarea fructelor, cu o influență reciprocă între ele (Tadiello et al., 2016a; Busatto și colab., 2017)., Pentru fructele climacterice, a crescut conținutul de IAA sunt necesare pentru a activa expresie a ACC sintetazei gene (ACS), codare ACC-sintetazei, care, la rândul său, va duce la producția de etilenă declanșând procesul de maturare (Figura 2C; Tatsuki et al., 2013). Fructe precum roșiile (Li et al., 2016) sau banane (Choudhury et al.,, 2008), care, de obicei, experiența redusă IAA conținutul înainte de debutul de maturare, arată o întârziere de maturare când IAA sau analogii sunt aplicate exogen, în timp ce fructe precum piersici, al cărui IAA conținutul crește progresiv până se coc pe deplin, arată maturare accelerată atunci când auxinele sunt aplicate (Tadiello et al., 2016b). Prin urmare, o reglare strânsă, complexă și diferențială a interacțiunilor auxin-etilenă trebuie să existe în diferite fructe în timpul maturării preharvest., În plus, IAA conținutul scădere înainte de coacere debutul în unele non-fructele climacterice a de-a reprima 9-cis-epoxycarotenoid dioxygenase (I) și începe ABA sinteza (Figura 2C; Jia et al., 2016). Interesant este că acest proces este, de asemenea, mediat de conjugarea IAA prin activitatea GH3-1 îmbunătățită (Böttcher et al., 2010). Cu toate acestea, Estrada-Johnson și colab. (2017) a arătat că, deși IAA conținutul reduce timpul de capsuni maturare, expresie a FaAux/IAA și FaARF familii genetice sunt induse în roșu recipiente, sugerând implicarea auxin de semnalizare în totalitate se coc fructele.,în timpul perioadei post-recoltare, conținutul de auxine rămâne invariant sau tinde să scadă datorită proceselor oxidative care pot determina reduceri mici, dar progresive, ale conținutului endogen al IAA (figura 2b). Auxin tratamente după recoltare întârziere supra-maturare în unele fructe (Chen et al., 2016; Moro și colab., 2017) și crește conținutul unor acizi organici, menținând aciditatea fructelor (Li et al., 2017), sugerând astfel că auxina joacă, de asemenea, un rol semnificativ în controlul maturării fructelor în perioada post-recoltare.,
mai Multe Roluri de Acid Salicilic în Timpul Dezvoltării Fructelor și Maturare
acidul Salicilic este un alt chorismate derivate fitohormoni care a fost în mare parte legate de efectul protector sub biotici de stres pentru a controla preharvest și post-recoltare a pierderilor de agent patogen fructe infecție (Babalar et al., 2007; Cao și colab., 2013). În general, conținutul endogen al SA liber este mai mare la începutul dezvoltării fructelor și apoi scade progresiv (Oikawa et al., 2015). Lu și colab. (2015) a raportat o creștere secundară a SA în timpul celei de-a doua faze de creștere a fructelor de piersici., Aplicarea exogenă a SA în banane a întârziat procesul de maturare reducând explozia respiratorie și raportul pulpă-coajă, precum și scăderea activității enzimelor legate de degradarea peretelui celular și a sistemului antioxidant (Srivastava și Dwivedi, 2000). Mai mult, tratamentul cireșilor dulci cu acid salicilic și acetilsalicilic (ASA) a sporit greutatea, fermitatea și culoarea cireșelor în stadiul comercial (Giménez et al., 2014). Aplicarea exogenă a SA sau a salicilatului de metil (MeSA) în timpul post-recoltare, întârzierea supra-maturării în fructe precum kiwi (Zhang et al.,, 2003) și cireșe dulci (Valero et al., 2011). Mai mult, Tratamentele cu salicilic sau ASA au atenuat leziunile de răcire ale rodiilor (Sayyari et al., 2011), tomatoes (Aghdam et al., 2014) și avocado (Glowacz et al., 2017). SA interacționează, de asemenea, cu etilena prin inhibarea expresiei ACC oxidază (ACO) (Shi și Zhang, 2012). În mod surprinzător, conținutul crescut de Expresie ACS în pere se găsește și după aplicarea SA într-o manieră reglementată de doză (Shi et al., 2013; Shi și Zhang, 2014). Prin urmare, reglarea producției de etilenă dependentă de SA este reglată fin (figura 2c).,
melatonina: un Regulator emergent pentru controlul dezvoltării fructelor
melatonina este, de asemenea, un hormon vegetal din calea derivată de corismat care a câștigat multă atenție în ultimii ani (Hernández-Ruiz et al., 2005; Arnao și Hernández-Ruiz, 2018; Sharif și colab., 2018). Recent, melatonina a fost evidențiată ca un factor important pentru setul de fructe, într-un mod similar efectelor IAA (figura 2a; Liu S. et al., 2018). În plus, a fost observată producția circadiană a acestei fitohormoni (Kolář et al., 1997)., În timpul maturării pe copaci, strugurii tratați cu melatonină au prezentat o sincronicitate mai mare și o greutate crescută atunci când se maturizează complet (Meng et al., 2015). Mai mult, un alt studiu recent realizat de Xu et al. (2018) a descris că melatonina ar putea promova maturarea boabelor de struguri prin interacțiunea sa cu ABA și etilenă, precum și peroxidul de hidrogen. S-a demonstrat că melatonina conferă toleranță la răcire în timpul depozitării la rece a fructelor, care pare a fi în mare parte legată de activitatea sa antioxidantă (Cao et al., 2018; Aghdam și colab., 2019)., În timpul perioadei post-recoltare, tomatele au prezentat o emisie crescută de etilenă după aplicarea melatoninei la 50 µM (Sun et al., 2015), spre deosebire de pere, unde aplicarea exogenă a melatoninei la 100 µM a inhibat producția de etilenă (Zhai et al., 2018), sau fructe de banane, unde melatonina la 50-200 µM a inhibat expresia ACS și ACO (figura 2c; Hu et al., 2017). Deși efectele diferite ale aplicării melatoninei pot fi legate de un efect de dozare, sunt necesare cercetări suplimentare pentru a înțelege mai bine rolul melatoninei în controlul dezvoltării fructelor.,
Cross-Talk de Chorismate Derivate Fitohormoni în Controlul de Fructe Dezvoltare
Hormonale diafonie între chorismate derivate fitohormoni a fost demonstrat de a apărea în timpul dezvoltării fructelor și maturare. Breitel și colab. (2016) a constatat că auxinele pot interacționa cu SA prin ARF2 (Figura 2C) și că supraexprimarea de ARF2 în tomate dus la scăderea conținutului de SA și o semnificativă întârziere de maturare., Acest lucru indică faptul că auxinele ar putea limita producția de SA în timpul dezvoltării fructelor, ceea ce ar putea explica un compromis între creșterea fructelor (mediată de auxin) și activarea apărării biotice (mediată de SA). Mai mult, aplicarea exogenă a SA pe fructele de papaya a dus la o expresie modificată a mai multor gene IAA, unele dintre ele fiind reglementate în jos, în timp ce altele reglementate în sus (Liu et al., 2017)., În acest caz, rezultatele au fost neconcludente de ce apare endogen în prezumtiv cross-talk între auxina și SA în timpul coacerii fructelor și sunt necesare cercetări suplimentare în acest și alte fructele climacterice. În orice caz, este clar că IAA și SA sunt strâns legate, nu numai metabolic, ci și funcțional. În cele din urmă, mai multe studii au raportat influența melatoninei asupra biosintezei și/sau semnalizării IAA și SA (revizuite în Arnao și Hernández-Ruiz, 2018), deși, din cunoștințele noastre, niciunul dintre aceste studii nu a fost efectuat nici în fructele climacterice, nici în fructele non-climacterice.,
Modulării Compușilor Bioactivi Biosinteza de Chorismate Derivate Fitohormoni
compuși Bioactivi (inclusiv compuși fenolici, isoprenoids, și vitamine antioxidante) nu doar au fost pe larg cercetate ca fiind responsabile pentru anumite proprietăți organoleptice a produselor alimentare, dar, de asemenea, pentru efectele lor protecție în celulele umane împotriva proceselor oxidative în dezvoltarea de neurodegenerative, boli cardiovasculare și anumite tipuri de cancer (Liu et al., 1999; Mueller și colab., 2010; Sturgeon și Ronnenberg, 2010; Björkman și colab., 2011)., Compuși fenolici contribui în mod semnificativ la conferind anumite arome, cum ar fi taninuri – responsabil de amărăciune sau astringenta gustul unor fructe, și culori, cum ar fi pigmenții antocianici – responsabil de roșu, albastru și violet culori de fructe (Croteau et al., 2000). Relevanța compușilor fenolici pentru consumul uman a fost asociat cu un efect protector împotriva proceselor oxidative în ceea ce privește cardiovascular și nervos central, sistemul de sanatate, precum si un risc redus de cancer al tractului gastro-intestinal (Björkman et al., 2011)., Carotenoide, tetraterpenes aparținând isoprenoids, joacă un rol în protecția împotriva foto-oxidativ, proceselor și ca pigmenți organici; ele sunt responsabile de culoare galben-portocaliu de fructe (Tapiero et al., 2004). Se consideră că carotenoizii dietetici oferă beneficii pentru sănătate în scăderea riscului de boli oculare și a anumitor tipuri de cancer datorită rolului lor de antioxidanți (Johnson, 2002). Vitaminele C și E, inclusiv acidul ascorbic și, respectiv, tocoferolii, sunt factori esențiali de calitate nutrițională în fructele cu multe activități biologice la om (Miret și Müller, 2017)., Având în vedere rolul relevant de toți acești compuși bioactivi ca promovarea sănătății compuși din fructe, modularea acumularea lor este de o importanță capitală în ambele climaterice și non-fructele climacterice (Vasconsuelo și Boland, 2007). Majoritatea acestor compuși bioactivi sunt acumulate la niveluri ridicate în timpul coacerii fructelor, când mai puțin gustoase fructe verzi conversa într-o nutritiv bogat, colorate și gustoase fructe (Daood et al., 1996; Ranalli și colab., 1998; Dumas și colab., 2003; Singh și colab., 2011)., Cu toate acestea, odată începută, maturarea nu poate fi blocată și, în general, duce la supra-maturare care afectează negativ calitatea fructelor (Kumar et al., 2014). Prin urmare, minimizarea alterării fructelor după recoltare, în timp ce obținerea unui conținut ridicat de compuși bioactivi rămâne una dintre cele mai mari provocări de rezolvat.auxina a fost identificată ca un regulator important al biosintezei carotenoidelor în timpul maturării fructelor de tomate climacterice. Maturarea în tomate este asociată cu degradarea clorofilei și cu trecerea xantofilelor la caroteni (β-caroten și licopen) (Fraser și colab., 1994)., IAA pare să întârzie tomate maturare prin reprimarea etilenă și mai multe în amonte de carotenoide transcrieri inclusiv Psy, a Ziso, Pds, Critiso precum și Chlases 1-3 și, pe de altă parte, promovează acumularea de β-Lyc1 și Crtr-β1 transcrieri, care rezultă în mai mare conținutul de xantofile și clorofila a (Su et al., 2015). În plus, în unele fructe non-climacterice, cum ar fi cireșele și boabele de struguri, auxina pare, de asemenea, să moduleze biosinteza antocianinei și, prin urmare, să controleze procesele de maturare a fructelor. Teribia și colab., (2016) a raportat o corelație negativă între conținutul IAA și conținutul de antocianină, indicând faptul că acumularea de antocianină începe atunci când conținutul IAA scade. Mai mult decât atât, aplicarea sintetice auxin-compus ca benzotiazol-2-oxyacetic acid (BTOA) a amânat până-regulamentul de gene care promovează enzimelor de biosinteza antocianilor, cum ar fi chalcone sintetazei și UDP-glucoză:flavonoide 3-O-glucoziltransferazei in struguri, fructe de padure (Davies et al., 1997).,s – a raportat că tratamentele Pre-și post-recoltare cu salicilați, inclusiv SA și derivații săi ASA și MeSA, reglează acumularea de compuși bioactivi care conduc la o activitate antioxidantă îmbunătățită atât în fructele climacterice, cât și în cele non-climacterice. Se știe că mai multe fructe, cum ar fi prunele, cireșele și caisele, au o viață scurtă, cu o deteriorare rapidă a calității după recoltare. Prin urmare, există o căutare constantă a tratamentelor care îmbunătățesc și mențin calitatea fructelor și, în special, conținutul de compuși bioactivi cu efecte benefice asupra sănătății., Tratamente de prune cu SA, ASA, si MeSA dus în semnificativ mai mare conținutul de acid ascorbic, antociani și compuși fenolici, atât la recoltare și după o lungă de depozitare la rece (40 de zile la 4°C; Martínez-Esplá et al., 2017). Rezultate similare ar putea fi observate pentru tratamentele preharvest cu SA,ASA și MeSA la cireșele dulci (Giménez et al., 2014, 2015, 2017). Mai mult, tratamentul post-recoltare Cu SA, ASA sau MeSA a menținut conținutul fenolic total, precum și conținutul de antocianină în timpul depozitării la rece în rodie (Sayyari et al., 2011), cireșe dulci (Valero și colab.,, 2011), cornelian cherry (Dokhanieh și colab., 2013), și caise (Wang et al., 2015). Aceste rezultate sugerează că salicilați poate fi implicată în activarea de fenilalanină amoniac liază, care este principala enzimă implicată în biosynthesic cale de compuși fenolici (Martínez-Esplá et al., 2017).recent, s-a demonstrat că melatonina reglează maturarea fructelor și modulează compușii bioactivi. Tratamentele Post-recoltare cu melatonină în fructele de piersici au crescut toleranța la răcire prin activarea sistemelor antioxidante., Refrigerare prejudiciu are loc în piersici fructe în timpul depozitare temperatură scăzută (între 2 și 7.6°C), caracterizată prin carne browning, tulburări de maturare și o sensibilitate mai mare la degradare (Lurie și Crisosto, 2005). Cao și colab. (2018) a raportat că, piersici tratate cu melatonină au aratat o mai mare transcriere abundenta de acid ascorbic biosinteza gene PpGME, PpGPP, și PpGLDH la a șaptea și PpGMPH la a 21-a și a 28-a zi de depozitare comparativ cu cel de control rezultând în creșterea acid ascorbic conținut., Mai mult, melatonina a indus creșteri ale activităților G6PDH, SKDH și PAL, care sunt enzimele esențiale pentru biosinteza compușilor fenolici. Autorii sugerează că tratamentul cu melatonină protejează fructele de piersici într-o anumită măsură de rănirea prin răcirea specifică a biosintezei compușilor fenolici (Gao et al., 2018). În plus, o analiză proteomică diferențială fără etichete a relevat efectul melatoninei asupra promovării maturării fructelor și acumulării de antociani după recoltare în fructele de roșii (Sun et al., 2016)., Autorii au raportat că melatonina exogenă a crescut de opt enzime legate de antociani cale inclusiv, inter alia, flavonol-3-hidroxilaza, flavanone 3 beta-hidroxilaza, anthocyanidin sintetazei/leucoanthocyanidin dioxygenase, și anthocyanidin 3-O-glucoziltransferazei. Mai mult, tratamentul post-recoltare cu melatonină a crescut fenolii și antocianinele totale în fructele de căpșuni (Aghdam și Fard, 2016) și a întârziat pierderea fenolilor, flavonoidelor și antocianinelor totale în fructele litchi (Zhang et al., 2018)., Tratamentele cu melatonină au crescut conținutul de fenoli, antociani și flavonoide din boabele de struguri (Xu et al., 2017). În plus, s-a raportat că melatonina sporește acumularea de licopen și producția de etilenă în tomate, sugerând că melatonina poate crește conținutul de licopen prin impactul biosintezei și semnalizării etilenei (Sun et al., 2015)., Participarea etilenei la reglarea indusă de melatonină a compușilor bioactivi a fost observată și în boabele de struguri, unde tratamentul dublu bloc al etilenei a arătat efecte reduse ale melatoninei asupra conținutului de polifenoli (Xu et al., 2017).se concluzionează că fitohormonii derivați de corismat, inclusiv auxina, SA și melatonina, nu numai că au un precursor comun, ci joacă roluri esențiale în reglarea creșterii și maturării fructelor., O discuție încrucișată metabolică și funcțională între ele și cu alți fitohormoni are loc într-o manieră spațiotemporală pentru a regla fin dezvoltarea fructelor climacterice și non-climacterice. Diferențele în dinamica fructelor climacterice și non-climacterice demonstrează că răspunsul hormonilor derivați de corismat nu este universal, ci mai degrabă specific speciilor. Mai mult, fitohormonii derivați de corismat modulează, de asemenea, acumularea de compuși bioactivi, influențând astfel calitatea fructelor., Suplimentare de cercetare este, totuși, nevoie pentru a înțelege mai bine cât de (i) altor hormoni, cum ar fi etilen-și ABA, modula lor biosinteza studierea metabolice cheie diversiune puncte, (ii) acestea interactioneaza la cea funcțională și moleculară niveluri, și (iii) au în comun modula compuși bioactivi biosinteza și, în consecință, influența nu numai fructe de creștere și maturare, dar, de asemenea, calitatea climaterice și non-climaterice fructe.SM-B a conceput și proiectat recenzia cu ajutorul MP-L. MP-L, PM, MM și SM-B au scris manuscrisul., MP-L a pregătit cifre cu ajutorul PM. Toți autorii au contribuit la discutarea ideilor, au revizuit și au aprobat manuscrisul final.această lucrare a fost finanțată printr-un premiu ICREA Academia acordat SM-B de către Guvernul Catalan.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Böttcher, C., Boss, PK, și Davies, C. (2011)., Preferințele substratului acil al unei IAA-amido sintetază reprezintă variații ale maturării strugurilor (Vitis vinifera L.) cauzate de diferiți compuși auxinici care indică importanța conjugării auxinelor în dezvoltarea plantelor. J. Exp. Bot. 62, 4267–4280. doi: 10.1093/jxb/err134
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Glowacz, M., Bill, M., Tinyane, P. P., și Sivakumar, D. (2017)., Menținerea calității post-recoltare a avocado „Hass” depozitat la rece prin modificarea conținutului și compoziției de acizi grași cu utilizarea compușilor volatili naturali – jasmonat de metil și salicilat de metil. J. Sci. Mâncare. Agric. 97, 5186–5193. doi: 10.1002 / jsfa.8400
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Zeng, W., și El, S. Y. (2010). Un rol important al flagellin receptorilor FLAGELLIN-SENSING2 în medierea stomatelor răspuns la Pseudomonas syringae pv tomate DC3000 în Arabidopsis. Physiol De Plante. 153, 1188–1198. doi: 10.1104 / PP. 110.,157016
PubMed Abstract | CrossRef Full Text | Google Scholar















